| |
EXPLANATORY ROUTES IN UNDERSTANDING EVOLUTIONARY TRANSITIONS INCLUDING THE ORIGIN OF LIFE
H.J. VAN DER MEER
There are lots of different theories, models, methods, and approaches, so it would be quite unfair to assert that physics or biology says this or that. It is just individual scientists who say this or that, and accordingly there is plenty of disagreement about what science as a whole says.
Michel Conrad, 1997
Abstract
During the evolution of the universe and life on earth, a number of transitions can be distinguished. One of these is abiogenesis, the transition from matter to life. Physical transitions relate to hadrons and atoms, molecular transitions to ion-bindings and polymers and organic transitions are about endosymbiosis and colonisation. As the transitions share some corresponding characteristics, they may be subject to the same mechanism. Varies approaches in understanding the underlying propulsion of evolutionary transitions are presented, like natural bonding, genesis of order, retro-causality and complexity. The universality of a transitional law and the presented hypotheses are mildly discussed.
Introduction
Our knowledge of the universe is limited. We have some ideas about its composition and origin, based on mathematical models, astrophysical observations and subatomic experiments with particle accelerators. We are confident that the entire universe is composed of the same material and that the known physical laws are independent of time and space, i.e. are universal. Accordingly, everything we find on earth we could find on any other planet in the universe, provided similar conditions (e.g. temperature, gravity, chemical composition). Even when the circumstances are quite different from our own, universal laws would still apply. One of those laws may concern evolutionary transitions.
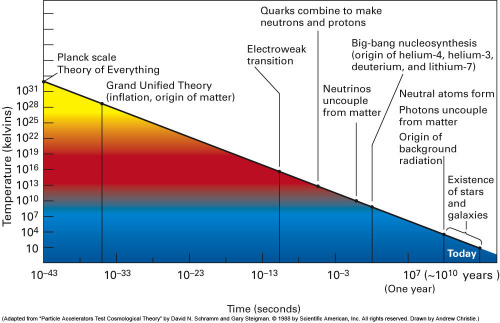
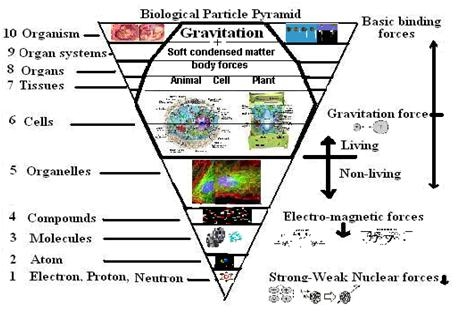
It must have struck many a biologist that from time to time developmental leaps occurred during organic evolution, but it was not until the mid-90’s of the preceding century that these transitions were seriously brought under the attention of the scientific community (Maynard Smith & Szathmáry, 1995). Their initial eight transitions include a number on the levels of replicating molecules and social behaviour. The list could be extended with new items like the development of protobionts out of chemical systems and the origin of the nervous system (Szathmáry & Fernando, 2011). Moreover, the organic transitions show a striking resemblance with the phase transitions during the origin of the universe with respect to hierarchal order (Weinberg, 1993). Accordingly, everything that exists seems to belong to one of a limited number of levels of organisation, from elementary particles to intelligent societies.
The hierarchal composition of the universe – from quarks and atoms via molecules to unicellular bacteria and neuronal networks – includes the transition from inanimate matter to living organisms (abiogenesis). It is generally accepted that life has arisen from inorganic matter, whether this happened on earth itself or had an extra-terrestrial origin. How abiogenesis actually can work is still far from clear and different theoretical models fight for recognition.
The most intriguing aspect of the sequent transitions is, that each intermediate phase encloses all previous ones and is itself part of the higher level(s) of organisation. What is it that makes such transitions come about? To answer this question, varies approaches are at hand, as described below.
Abiogenesis
The composition of Planet Earth may have changed in the course of its evolution, it has always been completely molecular. It is plausible to locate the origin of life within the region of the present biosphere, which is at the extreme periphery. It is also plausible to locate the origin of life within the environment that resembles the interior of present organisms, i.e. dominated by water molecules. As a medium, water-drops (damp or liquid) provide a range of manifestations of other chemicals.
Many a model has been developed and numerous experiments were carried out without convincingly point out how life on earth precisely originated. In the field of abiogenetic research one can distinguish three major questions:
- On locality. Did life originate on earth or elsewhere?
- On conditions. What were the conditions at the location where life originated?
- On the minimal claim. To what requirements must a protobiont come up?
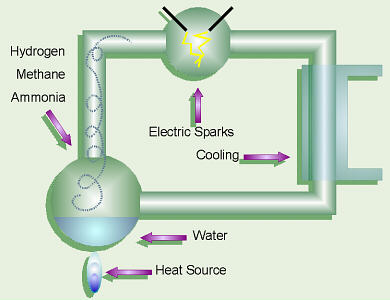
Many macromolecules that are essential for the origin of life have been synthesized under assumed prebiotic conditions on Earth (e.g. Parker et al., 2011; Martin & Russell, 2007; Fox, 1965), without any success so far to produce the slightest resemblance of a self-maintaining and replicating protobiont. So, the conditions and medium could as well be alien and the sympathy for extra-terrestrial origin of life is growing, as is the evidence (e.g. Nahle, 2003).
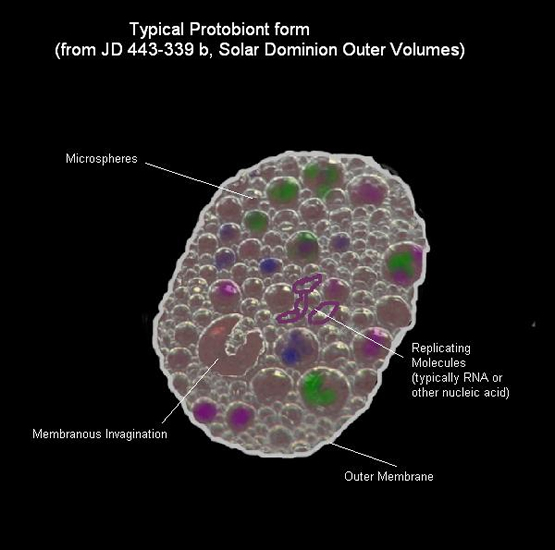
It is generally accepted that the minimum requirements of life are replication and metabolism. The combination of these two characteristics, together with an enveloping membrane according to some, are the necessary demands made upon an entity in order to consider it a protobiont. On the order of these characteristics (RNA world or metabolism first) is much dispute (Vasas et al., 2010). Besides, the lack of success in synthesizing a protobiont has led to otherwise creative approaches (Gibson et al., 2010).
Whether life has evolved on Earth or originated elsewhere (or a combination of both), abiogenesis must fulfil the same fundamental rules as all other evolutionary transitions of the cosmic hierarchy. Studying these transitions gives only little guarantee to answer the questions concerning abiogenesis. The reason that evolutionary transitions all seem to suffer the same synthetically unattainability may indicate that they can only occur in the complete absence of their own (and higher) levels of organisation.
Evolutionary transitions
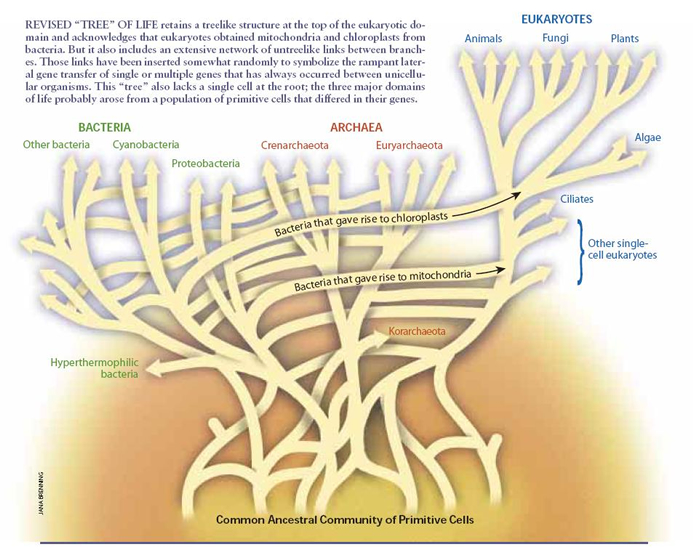
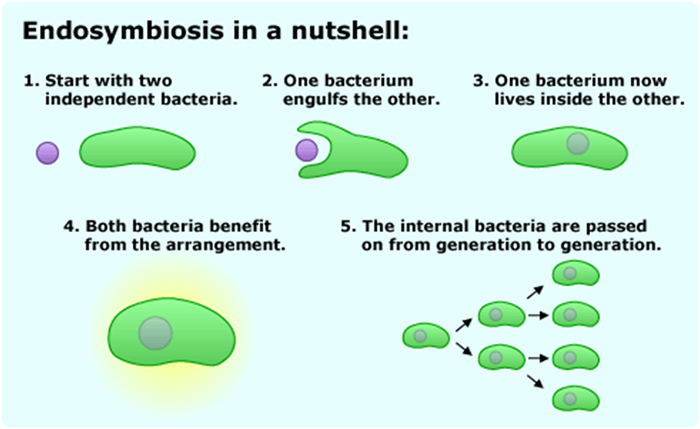
Central in the organic evolution on earth are the transition from the prokaryotes (bacteria) to the eukaryotic cell and the transition from single cellular protists to multicellular plants, fungi and animals (Szathmáry & Maynard Smith, 1995). A couple of other mentioned nexuses are the transitions from individuals to colonies and from social behaviour to cultural societies.
There are common features that recur in these transitions. Individual independent replication becomes an integrated part of a larger whole, an increased efficiency results from task specialization and the transitions show an abrupt increase of information flow. These characteristics may also be applied to the coming about of a global communication network.
The phase transitions that are assumed to have led to the cosmic building blocks involve quarks, hadrons and atoms of light elements like hydrogen and deuterium. As a result of initial quantum fluctuations, atoms clustered within galaxies and by gravitation into celestial bodies where they finally fused to heavier atoms. In the cooler vicinity of these stars, dust clouds of light elements and small molecular compounds accreted to form planets, like the earth. The increase of hierarchal organisation from quark to molecule is obvious and its resemblance with organic transitions undeniable.
Analogous to the organic hierarchy, one can draw a molecular hierarchy varying from simple chemical compounds to complexes of different polymers. Such a hierarchy should also account for the kind of bindings (ion-, covalent, etc.). Similar as in all hierarchal organisations, complex structures can exist by the grace of simple ones, suggesting an evolutionary development through several levels of molecular organisation. Obviously, the necessary transitions could not take place without the omnipresence of an appropriate medium and available molecules.
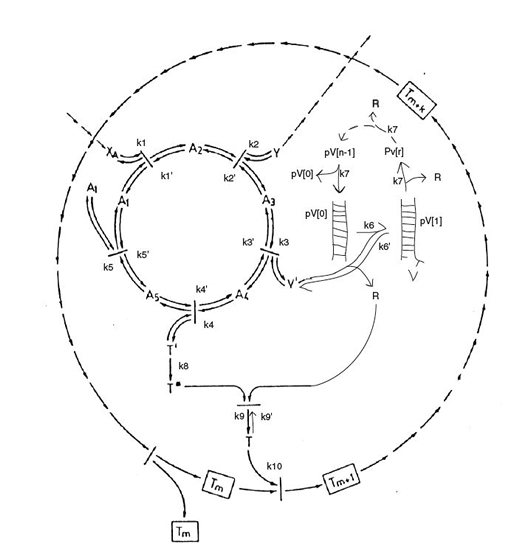
An inanimate structure may become alive when it comprises a metabolic engine, template replication and membrane formation (Griesemer & Szathmáry, 2009). All three are autocatalytic chemical systems that theoretically may have developed by autocatalytic polymerization. Their combination is less obvious. Yet, many combinations of still different chemical systems may have occurred with the aforementioned as the only one that survived.
Hypotheses/approaches
To search for a prevailing explanatory theory that would be applicable at all transitional levels must be a bit like chasing the rainbow. Yet, it is the ambition of many a scientific investigator to contribute to the search although most of them are dedicated followers of fashion. Therefore, a number of methodological directions of different popularity are presented below.
Natural bonding
In the last century the reductionist view of upward causation in physics and molecular biology has been extremely successful in understanding cosmic and organic evolution. It should not be a surprise to find that many a scientist persist in this approach to explore the hierarchal transitions.
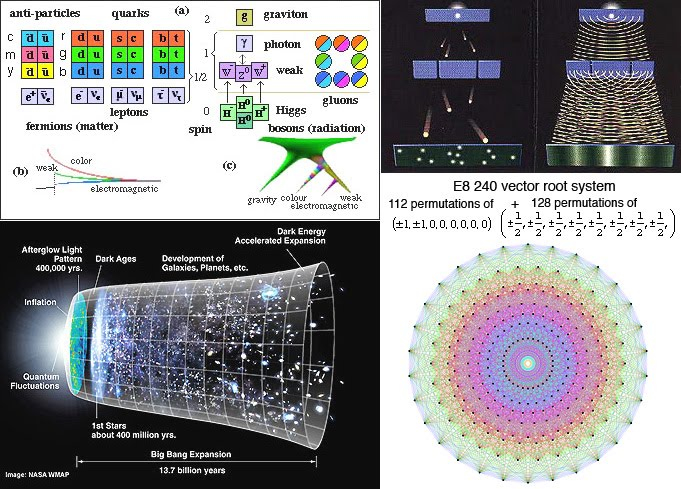
Fundamental are the smallest experimentally confirmed building blocks of matter, the up- and down quarks. According to quantum chromodynamics (QCD) the strong interaction mediated by gluons binds quarks together, in such a way that the mutual attraction increases with the mutual distance between quarks (Wilczek, 2006). The nuclear force, that binds protons and neutrons within the atomic nuclei, is understood as a residual effect of the strong interaction (Covello, 2003). Positively charged nuclei are neutralized by surrounding electrons. The drawing power between the nuclei, due to the residual effect, is largely restrained by the negatively charged atomic periphery and they cannot get any closer than the joined electrons in the forming of molecules permit. It should not be excluded that some residual attraction is left over to contribute – under specific circumstances, e.g. when neighbouring electrons reside in similar quantized energy states – to the formation of large and more complex molecules. Clearly, their coming about is mainly determined by electrostatic interaction, covalent bonding and van der Waals forces. The remaining non-linear interaction may give rise to a chain of cooperative weak-bonding effects, which enable the action of large protein enzymes (lysozyme with substrate centre) and the formation of supra-molecular complexes and cell organelles (Fielder & King, 2006).
The genesis of celestial bodies is ascribed to gravitation within high concentrations of atoms (the suggestion that gravitation may also be interpreted as a residue of the strong interaction remains mostly neglected; e.g. Lisi, 2007). Consequently light elements like hydrogen and helium are subject to fusion through which heavier elements originate. They form cosmic dust clouds in many regions of the galaxies and by gravitation the particles may accrete to one another in the emergence of planets like earth.
It is generally accepted that quantum mechanics play a prominent role in the coming about of chemical compounds (Atkins, 1991). The further enlargement of molecules should be understood in this context. The attractive ionic and covalent forces are merely effective over tiny distances which means that chemical compounds only come into being at sufficiently high concentrations of its components. As a consequence of quantum dynamics, these components are attracted to each other by van der Waals interaction as well. Next to this, the hydrogen bond too is a weak but essential attractive interaction between molecular components. In a molecular mixture with sufficient internal mobility (temperature) and variation and concentration of compounds, these forces – together and in due time – inevitably contribute to polymerization.
Life history on earth follows naturally from the basic interaction: novel attraction power emerged in cohesion with further molecular enlargement. Once autocatalytic replication presented itself, the phenomenon has spread with an exponential increase. As electron exchange can only occur when structures move closely, so is the transfer of energy: the input is not restricted to radiation, chemical compounds are necessary as well.
It has probably taken much longer before an enveloping membrane provided homeostasis for a population of replicating and energy transferring molecules that one could name a protobiont (Zimmer, 2009). To what measure the combination of components are determined by adhesion or mere coincidence is hard to say, once the protobiont is controlled from within by the storage and replication of information to do so, its existence and further development is assured. A constant need for input and production of output keeps chemical compounds together, by which concentrations of larger and smaller molecules are maintained.
During the initial abundance of input (food) many bionic forms emerge and competition increases as the input becomes more scarce. The organic interactions are dominated by exploitation (to consume and be consumed) while gradually cooperative interactions become more and more successful (Axelrod & Hamilton, 1981). Simultaneously, the competitive mechanism of natural selection shifts from an individual level to the level of groups (Thompson, 2000).
The organic transitions are each time characterized by the accumulation of entities that are held together by an increased efficiency of energy- and information transfer (e.g. division of labour; language). Following the above mentioned approach, all the cosmic and terrestrial phases of being can be reduced to the attractive interaction of the elementary building blocks and the transitions between them are the consequence of natural bonding.
Generating order
Entropy can be defined as the total of possible arrangements of agents within a system, without changing the system as a whole. It is a measure of the amount of disorder in a physical system. According to the second law of thermodynamics, the entropy of an isolated system which is not in equilibrium will tend to increase over time, approaching a maximum value at equilibrium. To the general opinion of modern astrophysicists, our universe expands with an exponentially increasing speed, implying that the universe should be regarded an isolated system, far from equilibrium. The lowest value of the cosmic entropy was at the beginning of the its expansion over thirteen billion years ago, and to some the maximum entropy is encountered in black holes (e.g. Greene, 2004).
Locally, however, a completely different picture can be drawn. Open systems within the ever expanding universe can develop order by lowering their entropy. This is accomplished by a constant flow of energy (matter) through the open system, in such a way that the total amount of entropy (including the energy source) increases. Living organisms are such open systems and their existence is due to this procedure. The generated entropy is dissipated out of the system in order to improve its own organisation at the expense of the environmental order (Nicolis & Prigogine, 1977).
Actually, the second law of thermodynamics says that, in a closed system, heat will not spontaneously flow from a colder body to a warmer one. This does not prevent increasing order in a practically closed system like our planet, as it receives low-entropy sunlight and reflects higher-entropy heat, which enables the coming about of local drops of entropy. Moreover, under specific circumstances, both entropy and order can increase (Adamson, 1998) and the gentle force of entropy may even produce order, e.g. in the hydrodynamic interactions of polymer molecules in aquatic solution (Hernandez-Ortiz et al., 2008).
Following the present approach, however, one operates a distinct schism between the inanimate and life. The inanimate universe, from quarks to quasars, tends to disorder, maximum entropy. At the same time and interrelated, living organisms evolved with an increasing amount of structural complexity and information flow, i.e. internal order. In other words, locally entropy decreases. The latter is permitted on the condition that the mean entropy increases. The genesis of order is therefor only possible in coherence with the genesis of disorder.
Consequently, primordial phase transitions, development of celestial bodies and chemical processes follow different rules than organic evolution does. And therefore a unifying law that holds for both inanimate and organic transitions still suffers of discrepancy. Besides, to understand the major transition of organic evolution alone, is hard enough (Calcott & Sterelny, 2011).
A different view of the universe is presented through the concept of wholeness and introducing two opposite kinds of order that are present throughout space-time (Giorbran, 2006; Bohm, 1980). According to this concept, physical reality emerges anywhere in the universe by a process of continuous unfolding and enfolding. Instead of order out of disorder, one kind of order emerges from the other kind and vice versa. The transitions in this view can be interpreted as different balancing stages between grouping order (explicate) and symmetry order (implicate). The holographic principle has given rise to a novel model of the universe as a nested hierarchy of information contents, each of which with a maximal limit of entropy density (the Bekenstein bound; Pesci, 2010). Extrapolating on the theory, a transition to the next level occurs every time that the limit of information content is reached (due to the maximal limit of entropy density). This should apply to all transitions, whether they concern inanimate, organic, social or ‘spiritual’ agents. The implicate order also means that it is impossible to gain complete understanding of a transition without understanding all the previous ones.
Syntropy and the future attractor
The duel solution of the second order Klein-Gordon equation suggests an opponent character: a positive solution of emitting waves and a negative one of resorbing waves (Cramer, 1986). Dirac’s relativistic energy/momentum/mass equation, introducing anti-matter, was supported by Anderson (1932) who empirically proofed the existence of negative energy. Instead of applying Pauli’s exclusion principle, one could also accept a symmetrical time flow: retarded waves from the past and advanced waves from the future. In other words, the duel solution of the KG equation suggests expansion associated with time moving forward and cohesion associated with time moving backwards (Di Corpo & Vannini, 2010).
Time-symmetry is also the central idea of the Wheeler-Feynman absorber theory (Wheeler & Feynman, 1945) introducing causality by advanced waves, presently called retrocausality. The concept of backward causation is supported by experiments in quantum mechanics (Kim et al., 2000) and the apparent time-paradox has been settled (Greenberger & Svozil, 2005).
Most scientists draw a distinct line between living organisms and inanimate matter, or, as Szent-Gyorgyi (1977) stated, “there is a basic difference between the living and non-living”. His postulated concept of syntropy, as an analogical counterpart of the principle of entropy, was eagerly adopted by creationists who recognized Aristoteles’ causa finalis as an argument pro irreducibility and against organic evolution. Yet, it is more elegant to associate syntropy with cohesion in a more universal sense, so that it can be applied to all phases of existence since the beginning of time. Syntropy, in this sense, should be regarded an organizing principle that, like entropy, can be attributed to the expanding universe and is constantly increasing.
The effect of syntropy can be described as the effect of a future attractor that gradually pulls everything to a higher level of organization. This is the case, not only in organic evolution, but also in the field of consciousness and global information-exchange at one end of the transitional spectre and in the genesis of inanimate matter at the other end. It is a controversial yet compelling approach to explain the coming about of all transitions.
Self-organising systems
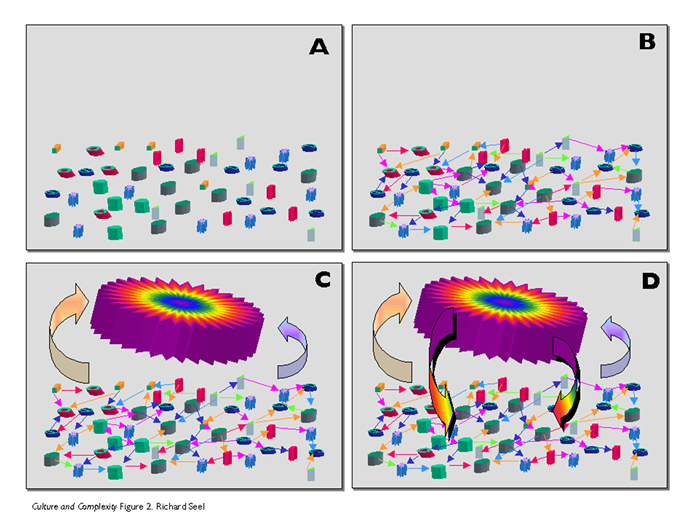
Somewhere in-between order and disorder we come across complexity. Complex systems are not regular nor random, they balance on the edge of chaos, which is a necessary condition for self-organisation (Kauffman, 1995). Lately, the inter-transitional phases are progressively more considered as complex systems with both upward and downward causation. The upward causation is locally determined by the mutual interactions among the component agents (each of which can be regarded a complex system itself). The downward causation or emergent behaviour of the system can be interpreted as whole-part causation or level entanglement and need not imply novel forces or laws (Davies, 2006). The network of local interactions operates as a non-linear process with initial random movements and finally stabilizing feedback loops, that gives the complex adaptive system its robustness and resilience (Heylighen, 2008). The more stable a system, the less controllable it is: after each perturbation it tends to return to its equilibrium state. It is the effect of feedback-loops that brings about the self-organisation and presents itself as the emergent property.
To understand the emergent behaviour it is necessary to realize that each agent tends to maximize its own fitness by mutual adaptation and coordination. The overall tendency towards a coherent stable state comes from the net effect of the positive and negative feedback-loops. The more similar the agents, the more dominating the positive feedback and the more regular and unified the emergent structure becomes. With more diverse agents the process is slowed down by an increase of negative feedback and the emergent structure is less predictable.
At some point, when the total of feedback loops reaches a dynamic equilibrium (stability), emergent properties appear as a resulting closure of the system. Many such closures may preserve and support the dynamic equilibrium, like autocatalysis by hypercycles (Hordijk et al., 2010) and an interfacing region limiting the system from its environment through which it can be conceived as a separate entity.
Discussion
Concepts on the history and construction of the universe are constantly under construction. Novel ideas to solve unanswered problems must have the potentials to be empirically tested. M-theory and the chaotic multiverse theory lack falsifiability and are therefore considered to be part of philosophy rather than of science. In time this may change. For instance, a two-dimensional universe has become less speculative since the holographic principle became more accepted after computations revealed that the maximal entropy in any region scales with the radius squared and not cubed (Stephens et al., 1994). Nevertheless, we are constantly confronted with paradoxes emerging from quantum reality, suggesting a fractal geometry of nature (Mandelbrot, 1982). The recurrence of phenomena on successive levels of organisation initiated the study of evolutionary transitions.
The postulation of a universal rule that holds for all transitions is hampered by their diversity. Apart from the varying number (from a few to innumerable) and divergent quality (similar/fraternal or different/egalitarian; Queller, 2000) of the nexuses’ components, umbrella theories suffer (i) hierarchal irregularities, (ii) include both open and closed systems and (iii) hardly convince while covering different, seemingly competing disciplines, each of which with its own laws and definitions (e.g. physical theory is described by rate-dependent dynamical laws that have no memory, while evolution depends, at least to some degree, on control of dynamics by rate-independent memory structures; Pattee, 2001).
The major difficulty seems to be the crossing between the inanimate and life. True to tradition, life on earth has always evoked a different approach than everything else in the universe, not in the least owing to the cultural context of civilisation which regarded life (humans) a holy entity and which seems to be very hard to break away from. The opposition that many novel views have encountered, also within the scientific community, shall never stop them to emerge.
There is no logical reason to deny a fundamental resemblance among the evolutionary transitions, including abiogenesis. The obvious resemblance they share is the increase of order. The primordial phase transitions coincide with an increasingly homogeneous distribution of particles in an expanding universe that continues throughout evolution. In aquatic solutions, different shaped molecules interact in such a way that some types tend to a homogeneous distribution (increased entropy) whereas other types form highly ordered constellations (Parades et al., 2009; Hamley, 2005; Cabodi et al., 2002). These concentration dependent interactions between polymer molecules may have played a prominent role in the transition from the inanimate polymers to a living protobiont.
The abovementioned approaches to understand transitions are not equally popular; some are even highly controversial. The combination of various flavours may yield a new recipe that gives satisfaction to all. Prigogine merged thermodynamics with general system theory to come to a better understanding of biological complexity and irreversibility (Prigogine & Stengers, 1984). The properties of specular holography with the use of laser light inspired many a scientist in their view on nature. In synergetics, self-organisation is mathematically explained as a reduction of the degrees of freedom (entropy) in a closed system by taking energy (cooling down) and in open systems by maintaining a flux of matter and energy through the system (Haken, 1983). David Bohm (1980) postulated that the ultimate nature of physical reality is an undivided whole in perpetual dynamic flux rather than a collection of separate objects. For Bohm, the insights of quantum mechanics and relativity theory point to a universe that is undivided and in which all parts merge and unite in one totality. He in turn inspired psychologist Karl Pribram to develop his holonomic brain theory (Pribram, 1991). After some period of vilification, the holographic principle has gained more general acceptance (Susskind, 2008).
The constructional concept of increasingly complex systems that evolved through space-time embraced both reductionist and holist views. Many studies on self-organising systems focussed on living organisms and their origin (Gánti, 2003; Kauffman, 1986; Varela et al., 1974) or artificial life (Holland & Melhuish, 2000; Clark, 1997; Langton, 1986). Lately, they concentrate on more general applications (Heylighen, 2010) and address systems at all descriptive levels (Jagers op Akkerhuis, 2010; Alvarez de Lorenzana, 2006). The latter studies pay dominant attention to the closure. The operator theory introduces the first-next possible closure (Jagers op Akkerhuis, 2008) implying a two-stage hierarchy of structural closures creating a physical boundary (e.g. electron shell or cell membrane) mediating a contained hypercyclic process (e.g. interacting molecules, prokaryotes or animal). As a consequence, the attention is focussed on the origin of cells and brains rather than on the transitions from prokaryotes to eukaryotes and from unicellular to multicellular organisms. On the other hand, the operator theory provides an inseparable transition from the inanimate to alive.
Another theory that links life to matter is provided by the fluctuon model (Conrad, 1998). Following a holistic view on quantum mechanics and applying the principle of philosophical relativity (Conrad, 1997), the theory explains the behaviour of macromolecules in living organisms as a manifesto of fluctuations in the sea of quantum vacuum. Rather impervious stuff which doesn’t mean that it makes no sense, just unknown, unloved. There are, probably, more theories connecting the topological structure of the vacuum with the measurable aspects of physical and biological events that are hardly known (e.g. Heim, 1994). Since complex systems involve both upward and downward causation, it is likely to hear, in the near future, of combinations between the abovementioned approaches. It should not be excluded that such combinations will include controversial, difficult and not-appealing theories as long as they fit in with the describing mathematics.
< click here to go back to PAN-OPTICS & EVOLUTION |
|
|