| |
Rapid Evolution (extended version of Dutch publication; Van der Meer, 2011)
Although most biologists are familiar with the principles of organic evolution, a brief survey of its major components seems sensible before presenting some examples of rapid evolutional processes.
Charles Darwin (1859) and Alfred Russell Wallace (1858) based their insight that organisms evolve by natural selection on four principles: first, that organisms have individual variations that are faithfully transmitted from parent to offspring; second, that all organisms produce more offspring than are required to replace themselves in the next generation; third that limited resources create a struggle for existence that regulates population size, such that most offspring die without reproducing; and fourth, that the individuals that survive and reproduce are, on average, by virtue of their individual variations, better suited to their local environment than those that do not (cited from Reznick & Ricklefs, 2009).
Such adaptive modifications within populations over time are now referred to as the products of microevolution.
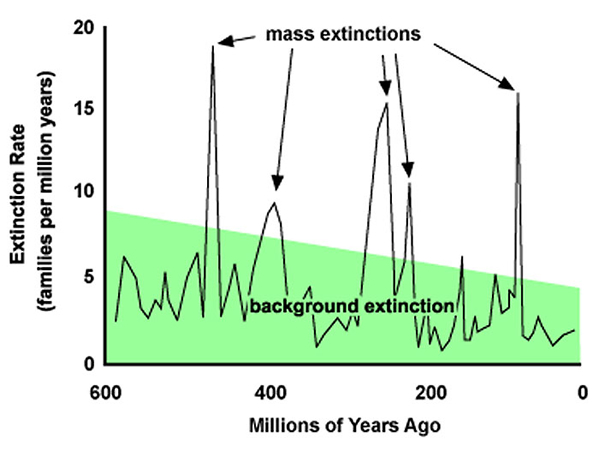
Macroevolution, by contrast, refers to the origin of new species and divisions of the taxonomic hierarchy above the species level, and also to the origin of complex adaptations, such as the vertebrate eye. In spite of the modern idea of extinction as primarily associated with modulations of the physical environment, the usual gaps that occur in the natural world are mainly the result of biotic interactions, like competition, predation, parasitism, and etcetera. In the struggle for existence, the strongest competitive interactions would be among closely related populations or within a population, because these organisms have the most similar requirements. Since these individual organisms cannot diverge from one another as they are part of an interbreeding gene pool, speciation also needs their reproductive isolation (Dobzhansky, 1937; Mayr, 1942).
Inspired by Sewell Wright’s adaptive landscape (Wright, 1932, 1982) that invoked the combination of natural selection, genetic drift, mutation and migration, allopatric speciation became the 20th century orthodoxy. It was generally believed that speciation could only occur when organisms became isolated by geographical barriers. Today, sympatric speciation is progressively accepted as equally important for the splitting of lineages: new species evolve as mate-choice is induced by behavioural differences or (sexual) preferences.
With the growing influence of molecular biology, organic evolution became increasingly conceived as primarily the result of genetic changes. Yet, the original concept of natural selection is almost completely based on ecological interactions that, occasionally, presents us with a clear demonstration of rapid evolution.
Reproductive parasitism
Inherited microorganisms that manipulate the reproduction of their host are a common feature in arthropod biology (Engelstädter & Hurst, 2009). Many insects are infected with Wolbachia, gram-negative bacteria that form intracellular inherited infections. Much of the success of Wolbachia can be attributed to the diverse phenotypes that result from infection. These range from classical mutualism to reproductive parasitism as characterized by the ability of Wolbachia to override chromosomal sex determination, induce parthenogenesis, selectively kill males, influence sperm competition and generate cytoplasmic incompatibility in early embryos (e.g. Bordenstein et al., 2010)
At the beginning of the millennium populations of the butterfly Hypolimnas bolina on the Samoan islands of Savaii and Upolu (South Pacific) were infected with the bacterium Wolbachia which killed male caterpillars. As a result most of the specimens in these populations were females that gave birth only to females. However, after approximately ten generations (about a year) the male population had evolved a gene that suppressed the bacterium and male numbers were up to about 39 percent of the overall population.
It is still unclear whether the gene that suppressed the parasite emerged from a mutation in the local population or whether it was introduced by migratory Southeast Asian butterflies in which the mutation already existed. However that may be, the findings constituted strong evidence that parasites can drive and substantially alter evolution (Charlat, et al., 2007).
Adaptive modification
One of the best known examples of a rapid evolutionary process is furnished by the peppered moth (Biston betularia) that is common in the British Isles. Among the species there are many individual differences of which the colouration of the wings is the most notable: some individuals are lightly coloured, others are dark.
Since the collecting of butterflies has been a favoured hobby in England for ages, the well preserved collections from the 19th and 20th century provided excellent material for scientific investigation. Around 1850 the interspecific colour variation was about the same as around 1950, with one striking difference. Among the moths of around 1950 there were many more dark specimens than light ones while among the moths of around 1850 it was just the other way around. Especially in the highly industrial area of the Midlands, the frequency of light moths was small in the middle of the 20th century (Kettlewell, 1955; criticized by Wells, 1999).
The moth flies at night and rests by day on tree trunks. It is preyed upon by birds that pluck it from its resting place by day. Before the industrialisation in the 19th century, the trunks of trees were encrusted with lichens. Against this background the light moth was practically invisible and the birds primarily picked out the darker ones.
In areas where air pollution became severe, the combination of toxic gases and soot had killed the lichens and blackened the trunks. Against such a background, the light form stood out sharply and the dark moth became abundant.
Dwarfism
Body size is of fundamental importance in understanding macroevolutionary patterns, both for extant taxa and for those with a fossil record (Gould & MacFadden, 2004). In the plant kingdom giantism is a rather common feature in many species as a result of multiple chromosomes but it can also be caused by evolving genes (Davis et al., 2007). In animals, ecological factors may promote dwarfism, at least in the Atlantic Salmon (Salmon salar) which then act as a powerful agent for selection and genetic differentiation (Johnston et al., 2005). Comparable reports on dwarfism (Levantine nanism) on other species are abundant (Sonin et al., 2007), although they may include the (indirect) effect of the physical environment.
Natural populations of guppies (Poecilia reticulate) in Trinidad are found in simple communities replicated in a series of drainages that can be characterized as either high or low predation (cited from Reznick et al.,.1997). Guppies from high-predation sites experience significantly higher mortality rates than those from low-predation sites. When moved to a low-predation site with further similar conditions, the population of guppies from a high-predation site attained within twenty generations maturity at later age and larger size. They also produced less and larger offspring than their ancestors on the high-predation site did.
The rate of evolution was up to seven orders of magnitude greater than rates inferred from the paleontological record. Apart from genetic recombination, these changes were probably also brought on by the postponed effects of predation on the relationship between population-size and available resources. Laboratory experiments, however, showed opposite results: the delayed senescence was observed in tanks to result from high predation, which is presumed to be caused by a decrease in the investment in the immune system (Reznick & Ghalambor, 2005).
Adaptive radiation
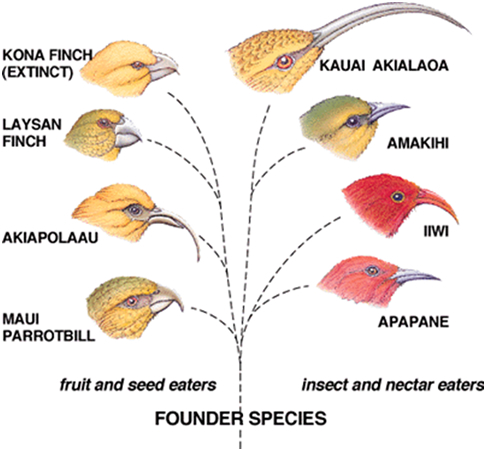
The most common situations which result in adaptive radiation occur following mass extinctions, or when species move into new, unoccupied regions (Losos & Ricklefs, 2009). The feature shared by these two situations is the availability of a variety of ecological niches. ‘Virgin’ regions are often islands and the involved colonists are endemic: they only occur in this one habitat. The Hawaiian silverswords (Asteraceae) are probably the foremost example of adaptive radiation among plants in the world. In animals the finches of the Galapagos are most certainly the best known species that came to be through adaptive radiation. It is a widely spread phenomenon and occurs among a variety of animal taxons, for example insects (Price, 2005), snails (Anderson, et al., 2010), reptiles (Rabosky & Glor, 2010) and mammals (Madsen , et al.,2001).
The African cichlid fish radiations are the most diverse extant animal radiations (Seehausen, 2006). Cichlid fish have radiated into endemic species assemblages in more than 30 African lakes and they are emerging as one of the potentially most powerful model systems in adaptive radiation research (Salzburger, 2009). Emerging experimental evidence suggests that they have undergone rapid speciation due to a combination of their molecular genetic potential and the influences of the environment on this potential. The genetic potential of the cichlids lies mainly in the ecomorphological plasticity of their feeding apparatus and their strong sexual selection (Chinsembu, 2009). Population genetic analyses confirm that the hundreds of haplochromine cichlid species in Lake Victoria evolved in about 10 to 15 thousand years from one or two ancestral species (Elmer et al., 2009).
Clearly, speciation rate declines through time as niches get filled up during the adaptive radiation and the number of species in cichlid radiations increased with lake size, supporting the prediction that species diversity increases with habitat heterogeneity and with opportunity for isolation by distance.
Introduced populations
Parallel to human traffic, intentional or otherwise, exotic organisms have invaded new environments. The difference with the adaptive radiations is, that instead of a broad diversification from one original species into many new ones, the modifications take place within one lineage. An example of exotic plants is for instance the native European St. John's wort (Hypericum perforatum) that was introduced in North America. The study on size, fecundity and leaf area suggests that introduced plants are evolving adaptations to broad-scale environmental conditions in their introduced range within a relative short period (Maron, et al., 2004). Introduced lizzards on an island in the Adriatic Sea evolved – within a couple of decades – increased bite strength and new structures in their digestive tracts together with a change in diet and population density (Herrel, et al., 2007).
The invaders may develop adaptive modifications themselves, their presence can also effect structures and/or behavior of the original species of the invaded habitat.
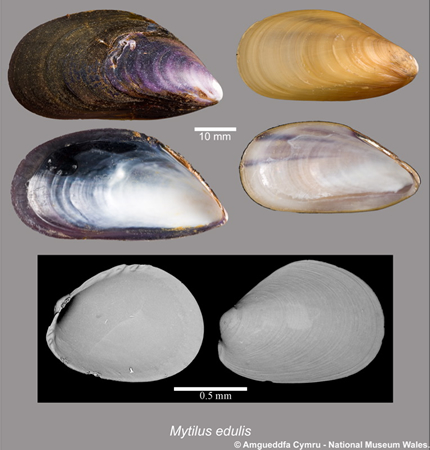
Since its arrival in the 1980s, the Asian shore crab (Carcinus maenas) has become one of the most dominant crab species in New England. But the area's native blue mussel (Mytlius edulis) has evolved to develop a defence against it in remarkably short order: when a mussel senses crabs nearby, it will thicken its shell; the result is a more secure mollusc – crabs usually crush mussels' shells (Freeman & Byers, 2006).
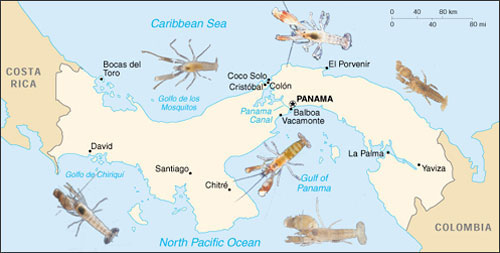
Recently more than 500 endemic haplochromine cichlids inhabited Lake Victoria alone even though the lake dried out completely less than 15.000 years ago (Witte et al., 2008). Most endemic cichlids of Lake Victoria disappeared after introduction of the Nile perch (Lates niloticus), which strongly muddled the ecosystem. As a result of intensive eutrophication the oxygen level dropped and water turbidity increased. After stabilisation of the population of the introduced predator, a number of haplochromine cichlid species appeared to have recovered. But not without a striking blow. In less than two decades these survivors had enlarged their habitat and fed on larger prey. They were also better equipped to visualize their surroundings with more adequate photoreception and to breath in their polluted environment with enlarged gills (Witte, et al., 2008). Most likely it is again the postponed effect of predation that sets the rapid evolutional changes. Close relatives with a narrow genetic margin disappeared to make room for the surviving species that could now occupy the vacant niches and habitats.
Sexual selection
Individual quality is often linked to appearance and the latter is generally believed to influence mate choice. This preference for congeners with specific features may eventually lead to sympatric speciation. Since the 20th century evolution-debate was dominated by discussions on allopatric speciation, sexual selection attracted serious scientific attention only a couple of decades ago. Sexual selection concerns the evolution of characteristics that foster the ability to acquire a mate of the opposite sex, either by mate choice (intersexual selection) or male-to-male competition (intrasexual selection). This kind of natural selection has only been investigated in animals with a focus on visual clues, although all senses could attribute to sexual selection. It was studied in for instance insects (Sirot, 2003), snails (Schilthuizen, et al., 2007), birds (Siefferman, & Hill, 2005; also often focussed on birdsong, e.g Catchpole, 1987), even mammals, mainly with respect to size-dimorphism (Schulte-Hostedde, et al., 2003).
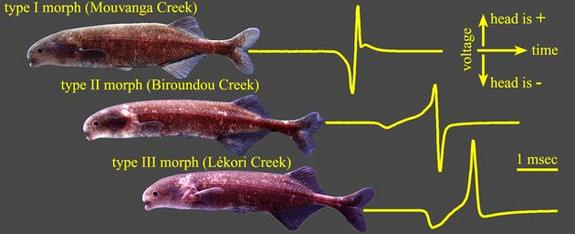
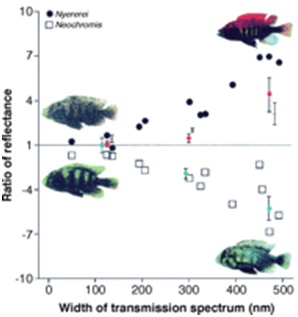
In Lake Victoria haplochromine cichlids inhabiting the shores of rocky islands hardly suffered the Nile perch predation during the second part of the last century. Among them were the species of the Pundamilia complex. These closely related species are brightly coloured and share a visual environment with a gradual shift of the penetrating light towards the longer wavelengths from top to bottom. The rather uniform coloured females are particular choosy in the selection of their mates. They prefer bright male nuptial colours over a paler appearance which correlates with the density of external parasites (Maan, et al., 2004; Seehausen & Van Alphen, 1999).
The shallow waters are dominated by the bluish P. pundamilia. Bright blue males have the widest appeal upon the females but they also experience the most competition among their own gender. Females of P. nyererei live slightly deeper and prefer the brightest red males. P. nyererei is slightly more sensitive for red light that is less intensely absorbed than short wave light by debris in the water (Carleton et al., 2005). And again, the brightest (red) males experience the most competition, in spite (or because) of their intersexual appeal.
It is suggested that the bright nuptial colours stand for higher fecundity as these males are less affected by parasites. On the other hand, brightly coloured specimens may be easier detected by predators. The specific habitats show much overlap and specific discrimination highly depends on the water transparency. The abovementioned process of sympatric speciation is undone by the recent eutrophication at the benefit of hybridization (Seehausen et al., 2008).
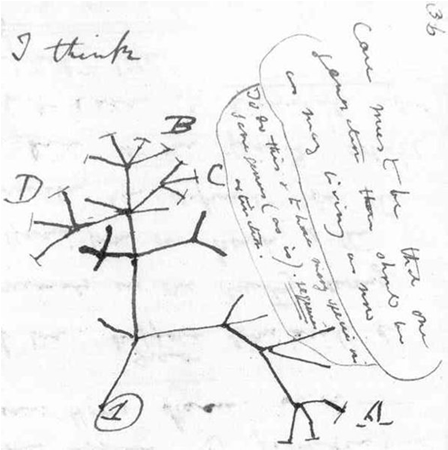
The abovementioned examples of rapid evolution do not present the principle of divergence as shown in a modern cladogram; they are in fact examples of microevolution. Only the Pundamilia complex may be an example of nascent speciation. Furthermore it should be realized that neo-darwinistic evolution can, by definition, only be applied to organisms with sexual reproduction.
Read more on speciation:
http://www.talkorigins.org/faqs/faq-speciation.html < click here to go back to PAN-OPTICS & EVOLUTION |
|
|