| |
Synergetic Evolution
To explain the immense variety of species, the mechanism of natural selection was introduced and defined in relation to sexual reproduction (Darwin, 1859). As a profound element of neo-Darwinian theory, natural selection became the major paradigm of all genetic evolution (Dobzhanski, 1937). For decennia natural selection was believed to work on the level of species and even ecosystems until it became clear that its strength was much greater on the level of the individual organism (Williams, 1957) and even stronger on the level of the genes (Williams, 1966; Dawkins, 1976). The mechanism of natural selection appeared to be most powerful on the genetic level to explain organic evolution without a purpose or future attractor. The evolution of altruistic behaviour could be understood in closely related specimens, although it is not yet beyond controversy (Foster et al., 2006). Natural selection appeared to favour cooperation between completely different species as was demonstrated with the theory of reciprocal altruism (Trivers, 1971) and game theory (Axelrod & Hamilton, 1981), leading to a better explanation of the evolution of altruism (Fletcher & Doebeli, 2008). Since the introduction of major transitions in evolution (Maynard Smith & Szathmáry, 1995), a resurgence of multilevel (group) selection appeared (Wilson et al., 2008; Michod, 1997) with promising potentials (Szolnoki & Perc, 2009; Bijma & Wade, 2008; O’Gorman, et al., 2008)
The philosophical objection to the basically reductional approach of selectionists has led to a proposed replacement of natural selection by for instance a theory of morphogenesis (Goodwin & Webster, 1996) or by natural experimentation (Reid, 2007). These contributions to the theory of evolution, that is still far from complete, are extremely valuable and call for special attention. However, I prefer their proposals next to natural selection rather than to replace natural selection.
Over three billion years of organic evolution show irrefutably the crucial role of concerted action and cooperation. The major transitions in evolution – or evolutionary nexuses – represent an increasing complexity that is based on cooperation and the conditions for its evolution has been listed (Nowak, 2006). It cannot be excluded that the few innovative events in organic evolution were the result of group selection rather than individual selection as they may be considered as the evolution of emergent properties of groups through differential group productivity mediated by trait-group inheritance (Thompson, 2000). At the beginning of each transition a new ‘virgin’ dimension became available to the emergent invaders who could rapidly radiate without competition (Koonin & Galperin, 2003) until natural selection forced them to adapt. Natural selection may not even be the primary source of biological organization, but a supporting actor that fine-tunes a self-organizing natural world (Kauffman, 1993). Yet, it is a compelling thought that group selection may, gradually, outbalances individual selection, each time a transition is at hand.
The transitions described below have some features in common. Each transition involves the junction of sub-divisions which provides an entity with a set of completely new attributes. As the entity or system is composed of the sub-divisions, it has increased in both complexity and size, at least with a factor 10. The transition can be interpreted as a step to a next level of organisation. In order to transfer, the involved components (agents) form a bond with sufficient fitness (Heylighen, 2000) to bound up the next level. The agents are basically not identical, they are often quite different (when many agents are involved they may occur in groups) and their original ‘freedom’ is transformed into a mutual alliance. It is not claimed that the transitions are equal, in a sense that they involve a related increase of complexity; they are mere remarkable events in the history of our universe. An appreciable aspect of the transitions is that they add something new rather than substitute something that already exists. Actually, the novel systems interact with the already existing levels of organisation, which provides yet an extra dimension of complexity.
Synergy
Haken (2004) used the term ‘synergetics’ to deal with systems, where order is an emergent property of macroscopic systems due to the interactions of elementary constituents (Érdi, 2008).
A synergetic process describes the development and sequence of emergent systems. A system is called emergent when the component agents provide innovative qualities that enables the system to ‘conquer a new dimension’. It is claimed that synergy played a key role in the development of complex systems, especially with respect to their co-operation (Corning, 1998). In (organic) evolution it may be regarded as a functional principle with respect to the fitness of emergent systems.
From a perspective of nonlinear dynamic systems models , synergy can be understood as the output of a self-organizing system with binary agents (subsystems) poising between a highly chaotic and a highly ordered state (Kauffman, 1991)
The processes and systems I will look upon are physical processes in the forming of elementary particles, atoms and molecules, chemical bonding in the origin of life, endosymbiosis in the coming about of eukaryotic cells, colonization and partial cell-division in forming multicellular organisms, landing by terrestrial symbionts, socialisation by individual organisms and global communication through electronic networks.
Synergy is often used in management consultancies to point out the advantages of cooperation. Frequently, these advantages are pointed out in defending the merger of companies, usually at the cost of workforce. In biological systems the term symbiosis is preferred when different species are involved. Synergy is also used as an umbrella term for irreducible cooperative effects of all kinds (Corning, 2003). This could easily induce misunderstanding as creationists believe that irreducible complexity affirms the intelligent design (Behe, 1996). I assume irreducibility to be illusive and that the incompetence to reduce complex structures is caused by dogmatic thinking and lack of creativity (see McDonald, 2002).
Non-linear oscillations may be unpredictable, once the picture becomes clear it is relatively simple to understand the causalities, by hindsight (Smale, 1980). So long as a causal explanation like natural selection enables the seeker to understand, there is no logical need to look for a divine designer (no doubt there will still be an esthetical and moral urge, but that is not the subject of this discourse).
Primordial Transitions
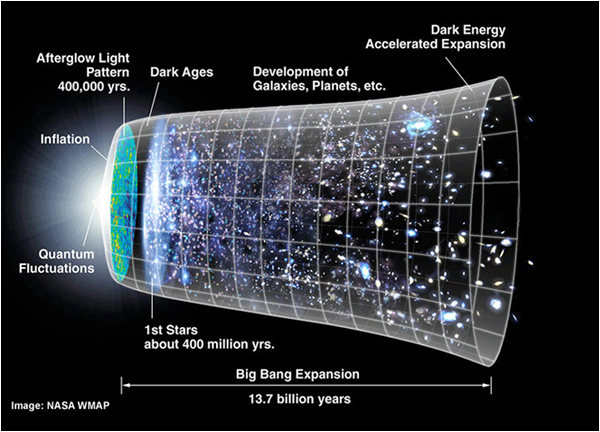
The organic evolution on earth was preceded by a much longer period of more than 10 billion years of inorganic evolution during which time our universe developed.
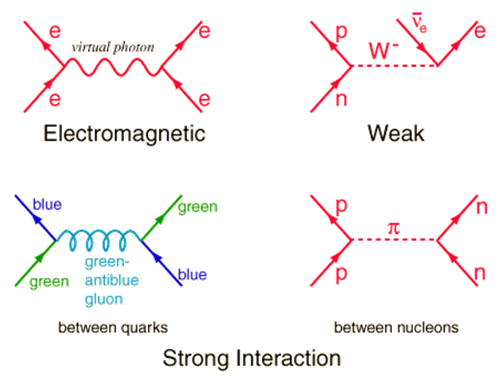
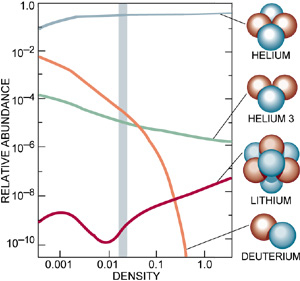
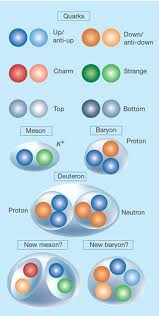
The mathematical descriptions of our universe are repeatedly under construction. Yet, astrophysical reports on the early development of matter and the later origin of galaxies and star-systems do agree on a starting point, when the universe was completely filled with energy and matter with infinite density (Gibson, 2005) and with slight quantum oscillations due to Heisenberg’s uncertainty principle (the latter caused local densities in a further homogenous space). From the first moment on, an immense hot universe expanded with great speed while the initial loose subatomic particles accreted in due time (Schewe & Stein, 2005). The primordial nucleosynthesis provided the initial synergic step in the forming of matter. Up- and down quarks joined together in forming protons and neutrons within a fraction of a second and after about three-quarter of the first million years most of the free electrons had been attracted to compose light element atoms (hydrogen, deuterium, helium and lithium). From that time on the universe became transparent (Ratra & Vogeley, 2008).
In the local denser regions of the ever expanding universe, gas clouds of light elements arose. Inside these clouds the density became so high (due to gravitation force) that stars and even black holes originated by fusion of the light elements. If the stars were big enough they would explode after fusion of all the lighter elements and throwing more heavy combustion-products into space. This process was repeated several times, when at last dust-clouds of elements (including heavier ones) circulating a star could evolve into planets, while the highly expanded universe had dropped to a few degrees above zero Kelvin (Greene, 2004; Hawking, 1988).
Entropy can be defined as the total of possible arrangements of agents within a system, without changing the system as a whole. It is a measure of the amount of chaos in a physical system. According to the second law of thermodynamics, the entropy of an isolated system which is not in equilibrium will tend to increase over time, approaching a maximum value at equilibrium. Ever since the beginning of time, however, the universe seems to be unable to regain its state of energy equilibrium as the expansion continuously accelerates (Frautschi, 1982; Krauss & Starkman, 2000; Watson, 2002; Lineweaver & Egan, 2008).The firm scientific opinion is, that the universe could hardly expand at all if it were not isolated from adjacent systems. Still, the system has been getting further and further away from an equilibrium distribution of energy and particles as the universal expansion continues to accelerate, thereby increasing the drive toward equilibration at the same time (Salthe & Furman, 2005).
Anyway, the synergic assemblage of elements from subatomic particles continued within the condensing intergalactic clouds, where densities increased to give birth to stars. According to the accretion theory (Safronov, 1969), planets could have evolved from the remaining circulating dust. Meanwhile a sufficient cooling down enabled various-sized atoms to combine and form molecules. This formation is conditional to catalyse further development anywhere in space. As much the largest part of the universe is hidden from any perception and even within our own galaxy this development is beyond our observation, we better don’t go far.
On our earth, for instance, the most common elements were iron and nitrogen. A heavy element like iron was drawn to the centre, while the atmosphere was mainly formed by gases escaping out of the cooling down of rocky materials. The presence of oxygen was immediately undone by oxidation of other elements like hydrogen, carbon and silicon, forming water, CO2 and quartz, respectively. Next to nitrogen, the early atmosphere must have had high concentrations of carbon-dioxide and vapour (H2O) as well as ammonia, hydrogen-sulphide and methane (Parker et al., 2011).
Due to mathematical difficulties, concepts of anti-particles and imaginary time were introduced (e.g. Hawking, 1982; Feynman, 1987).This procedure was preferred above the acceptance of a future cause as proposed by some, who, in contrast to the scientific establishment, follow a different reasoning. The ever growing complexity on a micro-level seems to be induced by an attractor that contradicts the increasing entropy (Mahulikar & Herwig, 2009).This negentropy or syntropy can be regarded an analogy of antimatter and is mathematically derived from the square root in the energy/momentum/mass equation that has both a positive (convergent) and negative (divergent) solution (Di Corpo & Vannini, 2010; Vannini, 2005). Syntropy is an intriguing concept that may induce complexity through strange attractors as in nonlinear dynamic systems (Stewart & Thompson, 1986). However, the concept is naturally embraced by creationists, making its use uncomfortable for scientific application. The second law of thermodynamics applies only to isolated systems, so – unless an ever expanding universe cannot be considered an isolated system – the use of syntropy is restricted to ‘open’ (energy-processing) systems, like living organisms.
Autocatalysis
The organic evolution on earth can be regarded as a continuation of the thermodynamic principle of energy transduction (Anilla & Anilla, 2008). Due to Heisenberg’s uncertainty principle and the Pauli exclusion principle, a range of molecules with their substantial characteristics could be produced by mere size-variation of different atoms and electron-exchange between them (Atkins, 1991). An immense variety of ‘free’ chemical compounds (today most of them are tied in some organic structure or process) enlarges the probability for the presence of catalysts to enable even more chemical structures. In a random mixture of reagents a catalyst will sooner or later be synthesized. This catalyst will direct further processes within the mixture by reducing the available compounds to enter into a random reaction. Initiating autocatalytic and oscillating reactions, the process becomes inevitable .
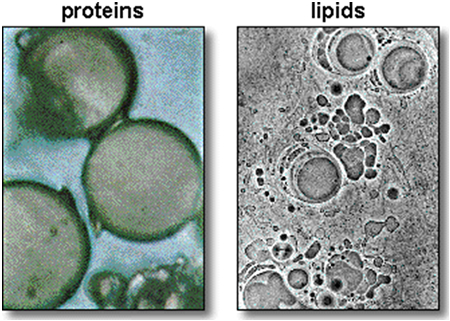
A reconstruction of the origin of life is highly hypothetical, yet there are some indisputable premises. To define an entity as a living thing, it should fulfil at least two conditions. The entity must be i) replicable in order to pass on information about itself, including the capacity to replicate, and the entity must ii) dispose of an internal environment separated from the outside world in order to generate energy (Monnard & Deamer, 2002; Peretó, 2005). Eventually, homeostasis could have come about – to some this may even be a preceding condition – by selective transudation of ions while the necessary energy was provided by chemical modulations in organic matter. The latter was abound by abiotic synthesis near volcanic eruptions (Johnson et al, 2008) and possibly by electrical discharges in the reductive gas atmosphere (Miller & Urey, 1959).
[In my opinion, viruses are not alive as they cannot generate energy on their own. Little is known about their early evolution but there is some evidence that they have contributed to the synergetic evolution by their inclusion in the genetic material of other organisms (e.g. Johnson, 2010).]
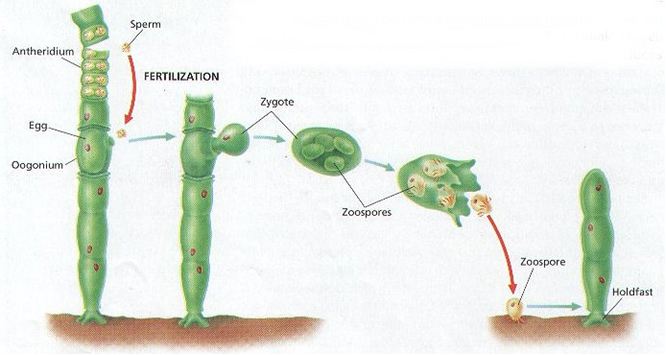
During the oceans of time for life to originate, replication needed only to happen once, probably with a substance like modern RNA (Nelson et al., 2000; Johnston et al., 2001; Joyce, 2002). By self- containment it would continue to replicate and develop further complexity to improve itself. This could explain why all modern organisms have the same structures to transfer information.
After the discovery of catalytic RNA’s (introns) among widely spread organisms (Lehman & Schmidt, 2003), there has been much speculation on their significance in early evolution of life (Penny, et al., 2009; Rodriguez-Trelles et al., 2006).
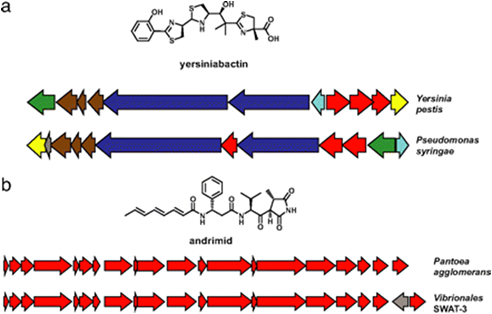
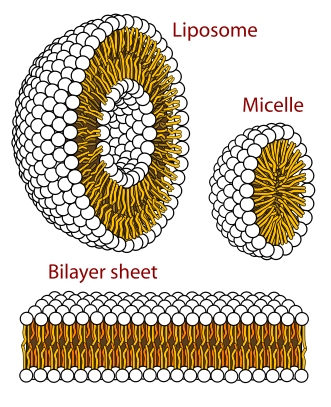
An autocatalytic chemical network seems a serious and plausible candidate to have produced the first RNA’s (Hordijk et al., 2010). This might have preceded by a reductive chemical reaction network to provide energy (as in modern chemotrophic bacteria). As a prebiotic metabolism, Sharov (2009) proposed an environment with autocatalytic non-polymeric molecules, like modern co-enzymes, as predecessors of nucleotides. These molecules would colonize the surface of self-assembling oil droplets, oxidizing the hydrocarbons in the oil and gradually transform the structure into a membrane. The disposal of an internal environment could also have otherwise been provided by self-organized lipid membranes (Segré, 2001; Trevors & Psenner, 2001; Morowitz, 1985).
The idea of self-organizing autocatalytic chemical systems as a basis for formation of early self-sustaining pre-metabolic molecular networks is seriously explored and has shown to be sufficient to allow for information transmission from one generation to the next (Monaco & Rateb de Motozon, 2005).
The primordial atmosphere was not only a highly reducing one by composition (primarily H2O, CO2 and N2), it mainly originated by impact degassing rather than coming from the Earth’s mantle itself (Zahnle et al., 2011). Quite in contrast with the long-term opinion that NH3 and CH4 were important components of the early atmosphere, these gasses would have only been present near volcanic eruptions and diluted in the oceans. The composition of the atmosphere is important for the origin of life regarding the available components in potential autocatalytic networks and regarding possible photochemical reactions and the sort of transmitted radiation from space.
The absence of ammonia and methane in the atmosphere actuated the hydrothermal hypothesis that suggests a submarine origin of life (Miller & Bada, 1993). And there is the theory of panspermia that presumes biochemical compounds have arrived from outer space (Hoyle & Wickramasinghe, 2000). Although an exogenic source of organic molecules to the origin of life is hampered by both the philosophical rejection of Ockham’s Razor and the more pragmatic consideration that only a small fraction would survive the impact (Pasek & Lauretta, 2008), it also gains in probability in an atmosphere generated by impact degassing. According to the opinion that evolutionary changes tend to accumulate gradually through a series of small steps, life started long before the formation of the earth and should therefore have originated on another planet (Sharov, 2010). His calculations, however, may suffer from false assumptions and insufficiently reckon with the attractors in the dynamical subsystems, which could reduce the period of accumulation substantially.
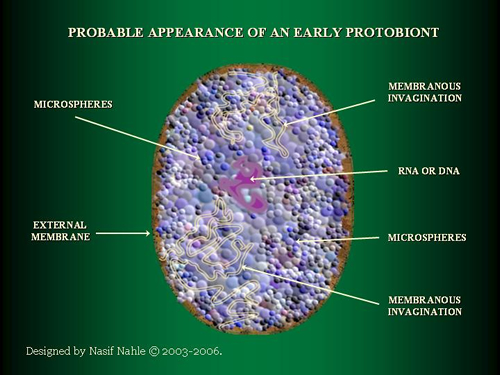
Among the numerous microspheres, some must have enclosed a replicating structure. The chance of a right combination may have been minute, the numbers of molecules and the available time were immense. My point is that the first complex structures that met the most fundamental demands of viability did evolve by fusion of correct ingredients rather than by selection. The properties of the separate components remain intact and their combination provides something that never was before. Perfect synergy.
Early Organic Evolution
Consider a single ancient microbe that replicates only once a year (if the environment is optimum, modern microbes may divide every 20 minutes) and consider its diameter to be 1 mm, then, after one century, the aquatic surface of the earth would be covered with a bacterial blanket of more than 3 mm thickness. You can calculate this yourself, with 2100 microbes with dimensions of 1 x 1 x 1 mm on a surface of 3,7 x 108 km2.
The first protobionts (a combination of an enveloping membrane, a replication apparatus and sufficient catalytic power to run the engine) were most likely chemotrophes with a reverse Krebs cycle and ‘consuming’ any available (in)organic matter. The energy generated by these autotrophic replicators would produce waste that sooner or later would be used in a similar way by other protobionts. And their waste again by others and so on. It is the beginning of organic co-evolution (interdependent relationship) and at the same time natural selection (competition) would make its biological debut. Autocatalytic replication of the separate building blocks became attuned to one another, in a way that is still totally unclear, and converted into reproduction.
The primordial soup of protobionts may have looked a bit chaotic, with many interactions, in- and exclusions, of large and tiny microspheres. Minimizing the number of membrane compounds would result in a spherical form due to physical law. Chemical properties of the compounds could have led to folds and elongation if the environmental pressure exceeded the morphogenetic costs, if any.
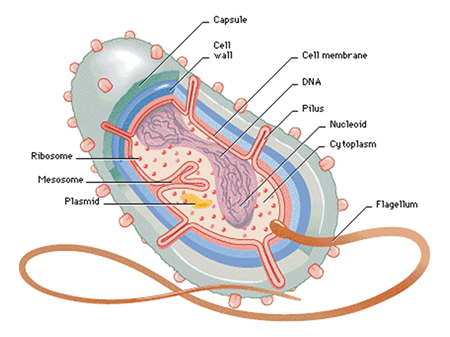
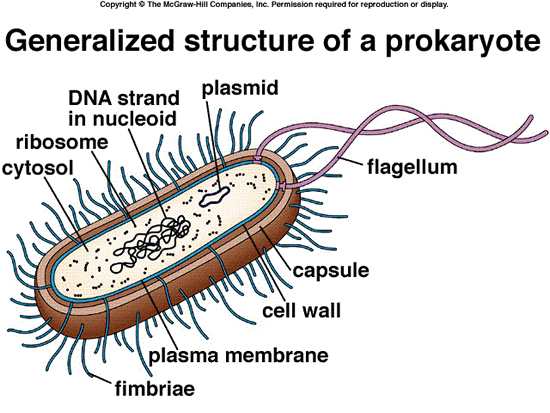
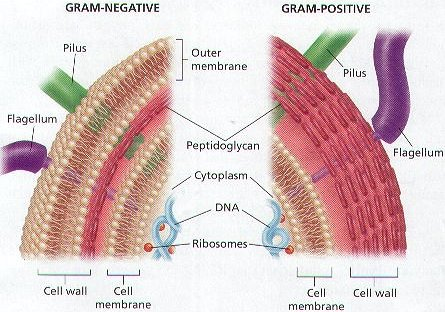
The earliest prokaryotic fossils – today the term prokaryotic is replaced by real bacterial – date about 3,5 billion years back (stromatolites). As the earth itself is about 4,5 billion years old, these prokaryotes must have evolved during the first billion years – the suggestion that living organisms invaded earth from outer space (Crick, 1981), only diverts the problem and unnecessarily complicates the matter (Sober, 1975). It seems plausible that many of the original life-forms disappeared as they were swallowed up, absorbed or included by more successful forms that were capable to ingest specific organic material in a more efficient way. This could be called natural selection, but not in the way Darwin defined it. However, one ancient group of bacteria, the Archeae do still exist and they stand out from all other modern life-forms by a specific composition of their membranes (Koga & Morii, 2007). The Archeae include the only modern life-forms that can survive extreme temperatures and salt-concentrations (extremophiles).
The modern bacteria are immensely numerous and can be classified in various ways. With respect to their energy-source the most common are photolithotrophes, that use light and an inorganic electron donor like H2O, H2S or S, and chemoorganotrophes, that use redox reactions and organic electron donors. To obtain carbon they can uptake CO2 (autotrophes like cyanobacteria)or receive it from other organisms (heterotrophes like parasites and decomposers). Regarding the electron acceptor the hetrotrophes can be either aerobe, using oxygen, or anaerobe, using a variety of (in)organic chemicals. Some of the present bacteria are aerobe but not autotrophic (nitrification bacteria), others use light but not CO2 (purple bacteria). Surely, their immense diversity must have been more trifling as modern bacteria highly depend on other life forms (and vice versa). Still, it was inevitably the matrix to further organic evolution.
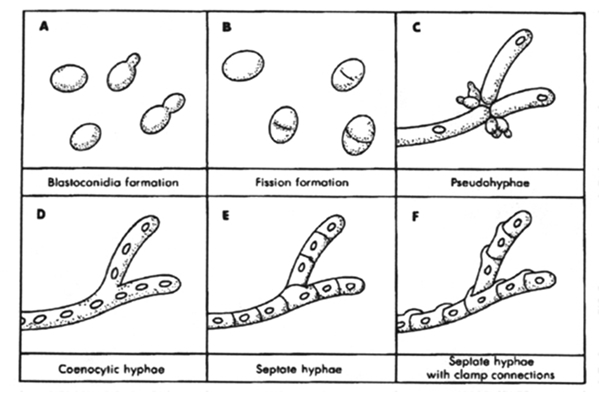
Reproduction of bacteria is primarily realized by binary fission while various ways of recombination are at hand. The unexpected frequently occurring lateral transfer of genetic material among various bacterial stems (Treangen & Rocha, 2011)does not say this happened three billion years ago as well, it still supports the idea of synergic evolution.
Taking the line that sufficient organic material was available to produce global growth, there is little doubt to my mind that the accumulation of microbes could easily have given rise to all sorts of fusion without neglecting the occurrence of selection by genetic mutations. There is no more direct proof for the occurrence of natural selection than there is for the occurrence of synergic evolution during the early billion years of organic evolution. It’s just a matter of philosophy.
Endosymbiosis
The real bacteria have cell membranes that are actually similar to those in all other organisms (apart from the Archeae). This fact as well as their simple structure (no organelles; free DNA) makes it plausible that all eukaryotes did evolve from the real bacteria. Among the many complex molecules some were radiation-absorbing proteins, through which electrons could be transferred to other molecules to generate some energy (Behe would argue that this could never have evolved stepwise whereupon McDonald would immediately prove him wrong). A primitive photosynthesis (finally as in modern cyanobacteria) replaced in some bacteria the chemical conversions that were necessary to generate energy. The by-product oxygen may have been quite harmful to many of the organic structures. It may have taken some time until the spin-off eventually could serve as an electron acceptor (finally as in modern nitrogen fixing bacteria).
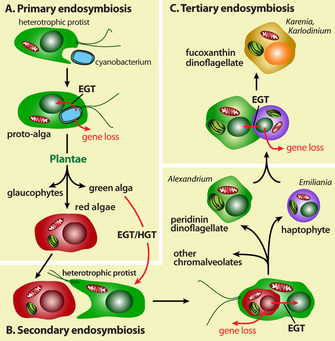
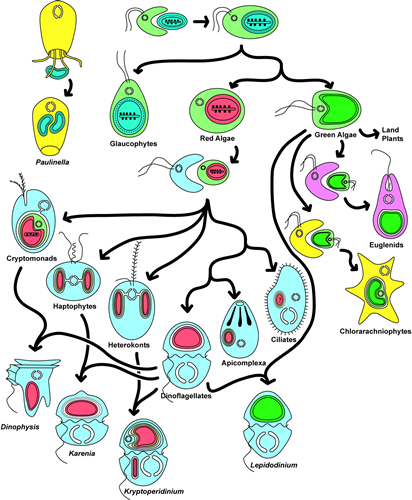
The remarkable similarities between the free living cyanobacteria and chloroplasts as seen in modern plant cells gave birth to the idea of symbiogenesis (Mehrezhkovsky, 1905). When also other organelles showed much conformity with free living bacteria and when they appeared to have DNA of their own, the serial-endosymbiotic theory was published (Sagan, 1967). Although not unanimously supported (e.g. Criswell, 2009) it is generally accepted that both chloroplasts and mitochondria have evolved from bacteria that were enclosed within a larger entity. The combination of photosynthesis and oxygen-recycling was another step in the synergic evolution. The basic substance of the eukaryotic cell, the nucleocytoplasm, has possibly developed out of the association between archeae and anaerobic heterotrophic bacteria while the nuclear envelop and endoplasmic reticulum came about by self-assemblage (Gupta & Golding, 1996). These so-called proto-eukaryotes would then have incorporated cyanobacteria and purple non-sulphur bacteria that we now know as chloroplasts and mitochondria respectively (Margulis, 1981). This hypothesis on a development of several billion years ago should be reviewed with some reservation. Contradicting theories exist on the origin of cilia and other movement-structures in eukaryotes. Advocates of the serial endosymbiotic theory argue that ancestors of modern spirochetes have penetrated into the proto-eukaryotes to form microtubules and undulipodia (Mitchell, 2007; Margulis et al., 2000). Endogenous models start with the premise that flagella arose from components of the eukaryotic cytoskeleton (Jékely, 2007; Cavalier-Smith, 1986).
On the origin of the eukaryotic nucleus the opinions are divided as well (Cavalier-Smith, 2010). The endogenous model is based on the presumption that the nucleus emerged selectively from a modified pre-existing endomembrane system (Jékely, 2008). According to the syntrophic model the nucleus originated from a fusion between archaea and bacteria (Lopez-Garcia & Moreira, 2006; Horiike, et al., 2001; Margulis et al., 2000)or at least from the endosymbiotic incorporation of archeael features into some host cell (Hartman et al., 2006). There is also an increasing interest in the possible role of viruses in the origin of the eukaryotes (Bell, 2009)
From Unicellular to Multicellular
Partial mitosis, sticking together after division, colony forming, the possible ways that multicellular organisms originated from single cells are various and limited. What they have in common is that co-operation between the cells is a necessity. Furusawa and Kaneko (2002) suggested that the emergence of multicellular organisms was a natural consequence of a cell colony that could grow continuously. All the aquatic organisms began their multicellularity by the products of cell division failing to separate, while most terrestrial microorganisms involve some form of motile aggregation
of cells or nuclei in a multinucleate syncytium (Bonner, 1998). Whether the multicellular organisms originated from aggregations of similar unicellular organisms as in modern slime molds, or that multicellularity arose from different species of unicellular organisms (symbiosis theory), in either case one could speak of synergic evolution. In closely related cells their co-operation can be understood from the ‘selfish gene’ point of view (Dawkins, 1976), whereas the formation of colonies of less related individual cells could be explained with the ‘prisoners dilemma’ (Heylighen, 1992) Since multicellularity probably evolved independently many times (Bonner, 2000), all possible scenario’s may have been realized.
Indissolubly connected to multicellularity are cell-differentiation and apoptosis (cell death) which may be rooted in their own history, that is the unicellular evolution (Ameisen, 2002). The increased genome complexity that coincides with multicellularity (Lynch & Conery, 2003) is most likely related to intercellular communication and the before mentioned processes.
Cooperative units of cells with different tasks would provide an enormous advantage compared to single cell organisms with respect to fecundity and viability. In unicellular individuals, the same cell must take care of both fitness components, whereas in multicellular organisms cells may specialize in one component or the other. A variety of selective pressures put a benefit on larger size and may push unicellular organisms to form groups(colonies) and evolve into multicellular individuals (Stanley, 1973). Large size can be beneficial for viability (e.g. in terms of predation avoidance, ability to catch bigger prey, a buffered environment within a group), as well as for fecundity (Bonner, 2000). As size increases further, the viability and fecundity benefits can be better achieved via the specialization of germ and the complete germ-soma separation; as a result, increased levels of complexity are achieved (Solari et al., 2003).
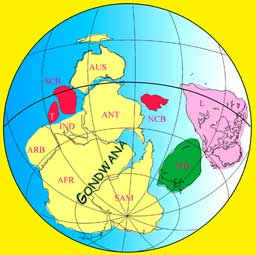
When multicellular organisms arose about half a billion years ago they exhibited many fundamentally different forms (body plans; Gould, 1991), including of all the modern phyla. The variation in body-plans remained limited as they were restricted by intrinsic rules of the subsystems during ontogeny (Webster & Goodwin, 1996). About 50 million years later, up to 85% of the multicellular organisms disappeared and most of the phyla were extinct forever. This first mass extinction that we know of, coincided with the forming of Gondwanaland that had drifted away from the equator. Apparently, the external conditions (shortened coastline and lowered mean temperature) worked as a selective mechanism in favour of a few body-plans over many, within complex ecosystems.
According to the fossil record, terrestrial plants and animals emerged relatively soon after multicellularity came into being (Pisani et al., 2004; Kenrick & Crane, 1997). As the oldest remnants of terrestrial organic life date over 2 billion years (Watanabe, 2000), it is plausible that the regarding microbial mats were available as nutrition for the later eukaryotic invaders. It is generally assumed that the first terrestrial eukaryotes could produce their own organic matter (autotrophs), in the course of which they contributed to a further increase of atmospheric oxygen levels. The terrestrial environment was quite hostile in the beginning, mainly because of high radiation levels as the ozone layer had not yet been completely developed to protect more complex life forms sufficiently. The land would have been nutrient poor, physically unstable and subject to violent fluctuations in temperature. Only lowland alluvial or lacustrine areas with fine-grained water- and nutrient-retentive soils may have been a suitable habitat for the first land plants of any size (Seldon & Edwards, 1989).
It is speculated that the colonization of land by eukaryotes probably was facilitated by a partnership (symbiosis) between a photosynthesizing organism (phototroph) and a fungus (Heckman et al., 2001). The fossil record in this respect is inadequate. However, the symbiotic partnership between fungi and algae or cyanobacteria already existed long before the evolution of vascular plants (Yuan et al., 2005) and their synergetic share in terrestrial invasion seems inevitable. With their hyphae the fungi could absorb water and dissolved inorganic ingredients while the enclosed phototrophs provided the necessary sugar. Photosynthesizing algae were much too vulnerable to survive on their own and were protected against too high amounts of UV-radiation and desiccation by the fungal cuticula.
Most modern embryophytes (land plants) still maintain a symbiotic relation with fungal hyphae as mycorrhiza to enlarge the contact-surface of the plant’s root system (Kiers, et al., 2011). Moreover, the mycorrhiza can provide nutrients that would be unobtainable for the plant without this cooperation.
Modern lichens consist of a symbiotic association of a fungus and a green alga. They can survive extreme conditions although some species are quite sensitive for air-pollution. Resent research revealed that bacteria as well may serve distinct functional roles within lichen symbioses (Bates et al., 2011).
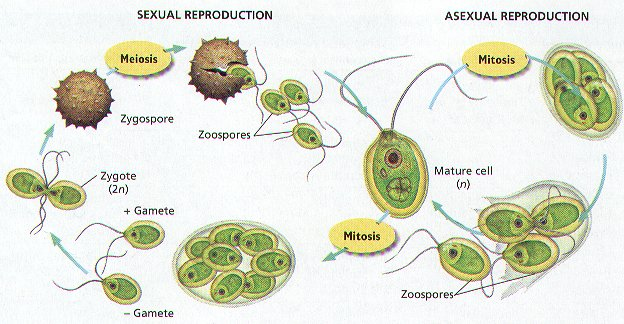
Various modern groups of algae are intermediates between unicellular and multicellualar plants as they form colonies. Best known among the green algae are the spherical colonies of Volvocales that are composed of numerous flagellates with occasional cytoplasmic interconnections and a distinct posterior (with red eyespots) and anterior. In a critical view on the transition from unicellular to multicellular, Calcott (2011) describes the evolution of multicellularity in Volvox carteri. Another green algae is the genus Scenedesmus that is chemically induced to form small nonmotile colonies, probably to prevent predation (the colony is too large to consume by zooplankton). Many golden and yellow-brown algae (chrysophyta) and diatoms (bacillarioophyceae) stick together in forming colonies, the latter with a rudimentary cell differentiation.
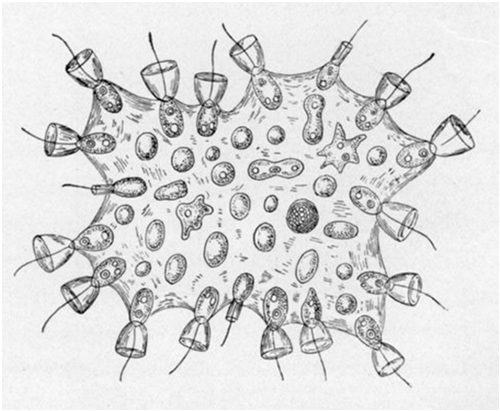
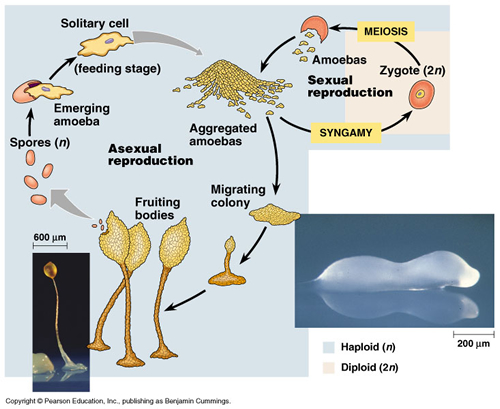
Modern slime molds begin as uninucleate amoeba and during growth only the nuclei divide and the organism develops into a large multinucleate plasmodium. When the conditions favour fruiting, the plasmodium breaks up into a single aggregates to form a fruiting body bearing haploid spores that give rise to the gametes of the next generation (Bonner, 1998).
The modern porifera (sponges) are multicellular organisms in-between single celled protozoa and metazoans with nervous systems and of all modern animals they share the closest relationship with the ancestral Urmetazoa. They are composed of a limited number of differentiated cells that, after separation, will re-join into the original animal. The existence of cell adhesion molecules allowed the emergence of a colonial organism (Müller, 2003). Sponges have no nervous and muscular tissue. Cellular communication and coordination is chemically generated by a complex set of molecular components rather than by electrical conduction (Meech, 2007).
Social Behaviour & Symbiosis
The occurrence of symbiosis, as an interdependent cooperation between different species, has already been mentioned above as clue-events of synergic evolution. Symbiosis is a source of evolutionary novelty and plays a crucial role in speciation and ecological diversification (Moran, 2006). The symbiotic relationship may involve multicellular organisms and microbes (this probably includes all fungi, plants and animals) that enable the more complex life form to survive, even under extreme conditions (Cordes et al., 2005).The organisms involved in a symbiosis may be fused to such degree that they cannot live apart. This may not always be the case in the mutually beneficial interactions between merely plants and animals. Nevertheless, the symbiotic network of their interactions play a prominent role in the functioning of an ecological system (Gomez et al., 2011; Bascompte & Jordano, 2007) and provides a higher level of complexity in its own. Synergy at work again. It need not be an effective force that is behind all this, the ascending steps on the ladder of synergetic evolution cannot be denied.
Among the eukaryotes, behaviour is usually attributed to animals exclusively. If the interaction between individuals of a species exceeds a certain amount of organization, one may speak of social behaviour. The definition is vague as there are many gradations of sociality and primarily the highest levels (eusociality, in a broad sense; Foster & Ratnieks, 2005) are worth mentioning in the context of synergic evolution. An exception is made for the cetacea. The completed reconquest of the hydrosphere by whales and dolphins favours its final success to the social aid of congeners during delivery of these mammals.
Emerging of individuated colonies occurred multiple times in many animal phyla (Wilson, 1975). Best known are the social insects but eusociality also occurs among shrimps and mole rats. For a long time evolutionists have mainly tried to understand how social behaviour (and its altruistic component) could have evolved from a selective point of view rather than along a synergetic approach and came up with kin selection and the selfish-gene-theory. Eusociality is more likely enforced by group selection than by individual selection and colonies are consistently superior to solitary competitors (Wilson & Hölldobler, 2005). Instead of molecular causes (like a gene for eusociality) ecological constraints seem to promote eusociality. The genetic relatedness may even be the result of enemy pressure as cooperation of individuals in constructing their complex nests and defending their highly valuable food-habitat resources benefits larger colonies over small ones (Reeve & Hölldobler, 2007). Eusociality seems above all the result of an unpredictable conjunction of circumstances. The valuable long-lived resource that fosters accumulation of closely related kin is scarce and therefor vulnerable to attack by competitors and /or predators and is worthwhile to defend. Strong selection from enemies on such kin-structured aggregations may promote evolution of specialized defenders that raise their own and the breeders’ inclusive fitness by defending the colony (cited from Duffy, 2002).
Apiculture already existed in ancient Egypt (Vogel, 1869) but it was not until the 1940’s that Von Frisch gained sight of their communicative movements (e.g. Grüter et al., 2008). It should not be surprising if more astonishing ways of communication were to be discovered in social insects in the future. The use of an analogy is misleading and often false. It is, nevertheless, irresistible to compare communicating ants with nervous cells and the ant-colony, as a sort of superorganism, with the nervous system (Hölldopler & Wilson, 2009; Gardner & Grafen, 2009; Hofstadter, 1979).
Symbiosis and eusociality come together when social animals exploit other organisms. Well-known are the cultivation of fungi and farming of aphids by some species of ants. Some ant species use other ant species as slaves and there are symbiotic relationships with plants, like acacia trees. Less known is the relationship between the eusocial shrimps and coral-reef sponges (Duffy, 2002). Of special interest is the symbiotic relationship between cockroaches and internal microbes that are claimed to have triggered eusociality in termites (Nalepa, 2011; Ohkuma et al., 2009). The mutual dependency between completely different organisms , at least one of which with its own complex eusociality, demonstrates a terrestrial biome, maybe even the entire biosphere, that reminds of the abovementioned microbe mats covering the entire earth, but then ever so much more complex. It is not at all surprising that this context of interacting inter-dependent living entities was conceived as an autopoietic system (Maturana, 1987) and inspired others to formulate the Gaia hypothesis (Lovelock, 1965; 2009).
Global Networks
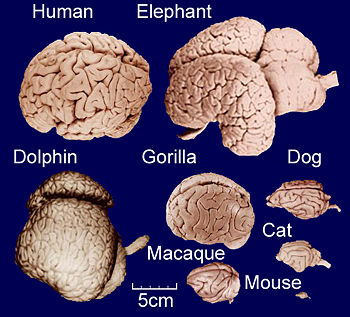
Humans have a fish-eye view with respect to life-history, and I am no exception. We play but a tiny role that we are apt to blow-up to proportions that may easily lead to misconception and disfigurement of a true perspective. Our species – and probably many other modern species – exists yet no longer than a couple of million years, which is, compared to all other living organisms, almost negligible in terms of time. Our brain seems nothing more than a protuberance of nervous tissue. We experience it, however, as the residence of our cognition and (self)consciousness. In the present context (self)consciousness can be regarded a spin-off. We share the larger part of our brain with other vertebrates mainly for the purpose of perception and to reflect on sensations (Humphrey, 2011). On top of that, our large frontal lobes are primarily used in data processing, a feature that could only have developed so well in an extreme social being (Dennet, 1991). The powerful capacity of data processing enabled Homo sapiens to develop a major innovation: the unequalled manipulation of the mineral world yielded two reluctant steps on the ascending (to some an uncomfortable predicate as they label it with value) ladder of synergic evolution, namely the primordial development of space travel and cyberspace. Critics of the Gaya hypothesis, in which the biosphere is presented as a sort of super-organism, pointed at the impotence of the Earth to reproduce. This was already disproved by the suggestion of humans potential to explore space and colonize other planets (Sagan & Agel, 1973).
The global information network that our species constructed, provides another step in synergetic evolution. Today, practically every human being can communicate with everybody else on this planet. Within seconds information that might be important for the maintenance or improvement of smaller or larger units of society can be spread to every corner of the earth. Of course, much of the information is irrelevant and even false, as well as this is the case in the individual brain and in DNA. That does not alter the synergetic impact of cyberspace in everyday life.
Not only information matters. There is also an elegant flow of economic interdependency meandering the planet, in order to provide the necessary energy for the maintenance of social communities (often at the cost of others). The major force to maintain this process is a universal urge of smaller units, even individuals, to extend. And last but not least, there is a fundamental human need for security, possibly an essential of eusociality, that drives us together even in disagree.
Today there is also much concern for our environment. Many people fear that organisms of all sorts suffer extinction by our expanding dominance. Some of the present organisms are directly needed for our own preservation and others are, by the netted relationships as described above, indirectly needed. We have descended from earlier organic developments, but so have many other present living things. And many of them are actually dead ends. Most people have no problems with exterminating weeds and bugs, but it is an interesting issue when projects are suspended for the benefit of a harmless but rare species. Still, our mere existence is extremely interconnected with the ‘rest of the world’.
We find it self-evident that everything is constructed out of atoms and molecules and we take it for granted that these tiny building blocks are present as individual materials as well. We experience it as less self-evident that the 1030 bacteria on earth comprise half of the earth’s biomass. And without them the eukaryotic cell would never have evolved. There are still at least a quarter of a million unicellular eukaryotic species, in spite of, or maybe thanks to, the origin of plants, animals and fungi. Despite the suffering from many parasites, multicellular organisms may never have evolved without the already present bacteria and unicellular eukaryotes. Eusociality and human social behaviour in the presence of the before mentioned life-forms were perhaps even reinforced by some (in agriculture, cattle-breeding and sanitation). The fitness of each organisms highly depends on both its success to consume other organisms and to avoid its own dispatch. In the context of major transitions, the similarity between humans and protobionts is that both use inanimate matter to extend (by replication, reproduction, communication and extra-terrestrial exploration). The global network is not restricted to a silicon-based information network, it is also the total of layered organic interactions (the top ones usually referred to as ecosystems) as well as it is subject to the laws of participation in our solar system, our galaxy, forming a part of the non-local universe (Aspect, 2007; Fiscaletti & Sorli, 2007).
The Future
It is generally accepted that evolutionary processes are more or less arbitrary. If the process was restarted with all the similar conditions, the outcome would in detail be quite different. I do agree with the opinion that the origin of species highly depends on accidental circumstances, a natural development that allows no repetition. Concerning the primal transitions, however, I am convinced of a fundamental law by which anywhere in the time-space continuum, given sufficient time, elegant systems fuse to enter a new level of complexity. This philosophy enables me not only to look backwards but also to make predictions about the future. On earth, the transitions from molecules to the descend of man involved a stepwise 109 x size increase (10-10 – 10-1) which took about 4,5 billion years. If we consider the global network as realized today, there has been a 108 x size increase (10-1 – 107)since the origin of hominids about 5 million years ago. Accordingly, it would take no more than another millennium before our solar system (1012) has been colonized.
The optimistic view on human progress predicts an exponential growth of sustainable technology and cybernetic developments through which finally all ecological and social-cultural problems in the world will be solved (probably restricted to a happy few). Out of the global information network and future technological innovations, another transition will emerge: Homo sapiens will give the first impulse to a new sort of evolution, namely the development of self-assembling robots that will colonize the galaxy.
Pessimists do not believe that ’human greed will be able to cope with Gaia’s revenge’. They prophecy the downfall of human civilisation for the benefit of small-scale nature and they fear a final extinction of Homo sapiens. If human species is indeed a dead end in organic evolution, a completely different path may be followed. A probable transition would have been the combination of behavioural traits (animals) and photosynthesis (plants) in order to develop eusocial animals that no longer depend on organic compounds to survive. This, of course, is fiction and it would probably take hundreds of millions of years for such organisms to evolve.
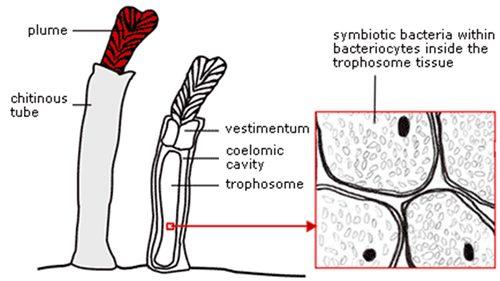
Yet today, various forms of symbiosis exist between animals and taxa of other kingdoms to reduce organic dependency. Chemoautotrophic bacteria live in a symbiotic relationship with the giant tube worm Riftia pachyptila . By the oxidation of H2S from the ‘black smokers’ on the ocean floor along the Galapagos rift, these bacteria provide the worms with the necessary energy to synthetize organic compounds (Van Dover, 1996). The details of the symbiotic process are yet unclear. Other organisms, like clams and mussels, use similar symbiotic mechanisms to obtain their nutrition. Some animals maintain a symbiotic relationship with algae that offer oxygen produced by photosynthesis in return of nitrogen waste (Kerney et al., 2011). The sea slug Elysia chlorotica has even obtained photosynthetic power itself, by incorporating algal chloroplasts inside the epithelium cells of its digestive system and by providing the essential plastid proteins by horizontal gene transfer (Rumpho et al., 2008).
A completed conquest of the atmosphere would be another long-term evolutionary challenge. Beyond question the air is full of tiny organisms, predominantly bacteria. Super-flying eukaryotes have only evolved among insects and birds. In insects, the ability to fly does not come about until the last stage of metamorphosis, i.e. in the adult specimens, and is at present primarily intended to find a mate for sexual reproduction as feeding occurs during the earlier stages of growth. These animals remain bound to earth in their dependence of nutriments on the ground, like e.g. leaves, blood or nectar. Many birds do not have that problem as they feed on aerial insects. At present, several species of birds, like the swift Apus apus , can stay in the air for months and even sleeps while flying (Rattenborg, 2006). Given time, these birds may become completely air-borne, in a sense that they carry their eggs and young while flying. The chance of success lies mainly in the readiness to cooperate, in this case with congeners.
Still, I support the optimistic view in the understanding that there is still a long way to go before the transition from human beings as social animals to a uniform and stable global society has become fact. And the outcome of the process, how the world will look like then, is, as in any evolutionary development, quite unpredictable.
< click here to go back to PAN-OPTICS & EVOLUTION |
|
|