| |
Introduction
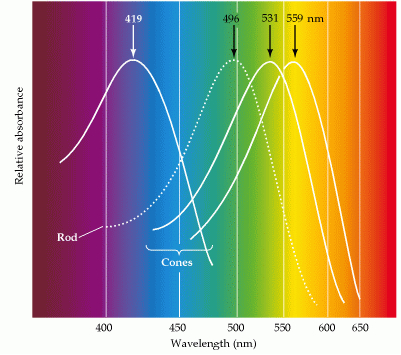
Duplex vision is a common feature of the vertebrate eye; it means that the vertebrate retina has two separate systems to detect light (Schultze, 1866; Von Frisch, 1925; Walls, 1942). The scotopic system is only used under dim light conditions and is generated by rods, all of which contain the same rhodopsine photopigment. The photopic system becomes activated when more light is available and is generated by usually two or more different cone-types containing different photopigments, which enables the system to distinguish between different wavelengths (colour vision; Bowmaker, 1991; Jacobs, 1993; Harosi, 1994). The two systems interact with the retinal pigment epithelium to maintain visual functions (Strauss, 2004) resulting in e.g. circadian regulation of retinomotor movements in lower vertebrates (Ali, 1975; Pierce & Besharse, 1985)
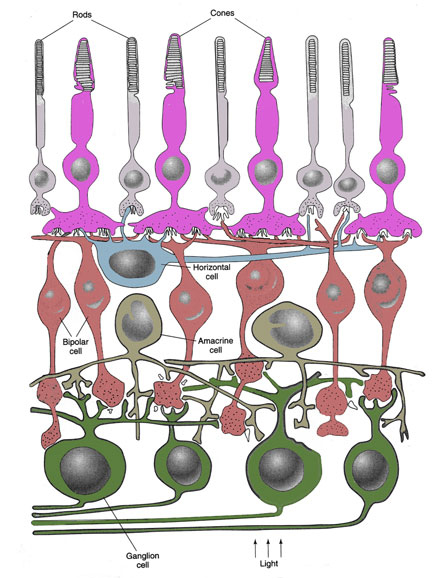
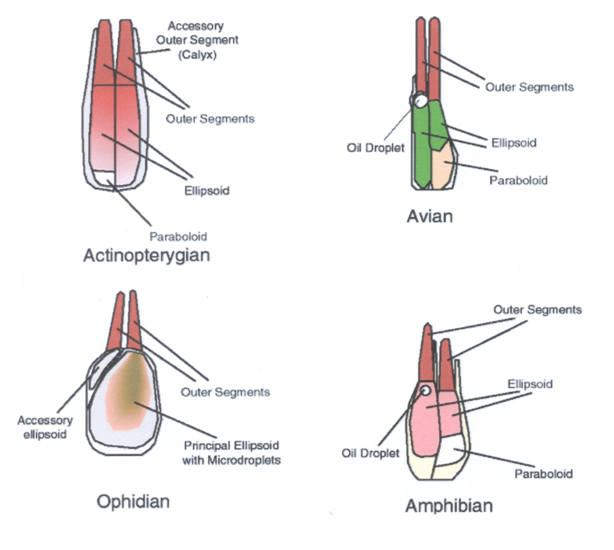
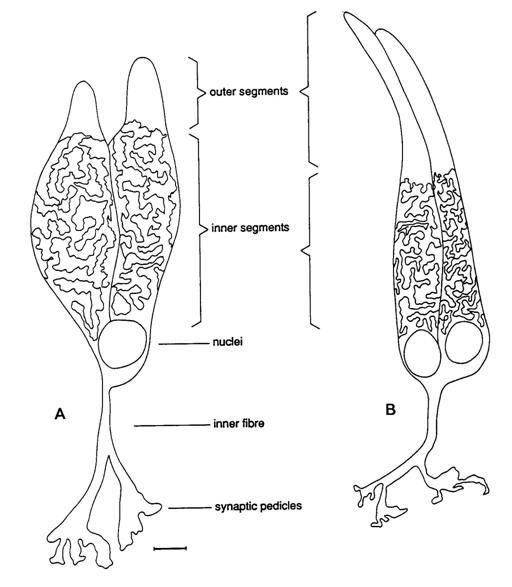
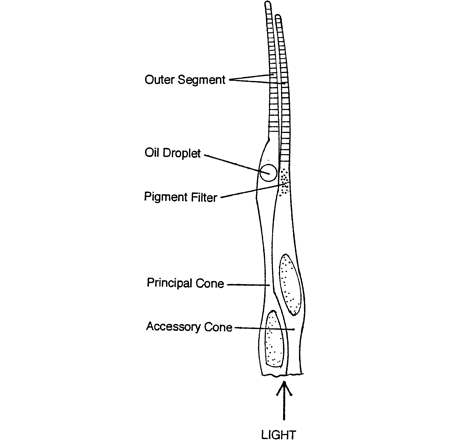
The photoreceptors are modified nerve cells with distinguishable compartments. The vitread positioned cell body containing the nucleus has many synaptic contacts with neighbouring photoreceptors, horizontal- and bipolar cells, the sclerad positioned outer segment containing photopigments absorbs light of the photopigment matching wavelength and in between these two compartments one can find the ellipsoid or inner segment containing many mitochondria that provides the necessary energy (e.g. Kolb et al., 2001)
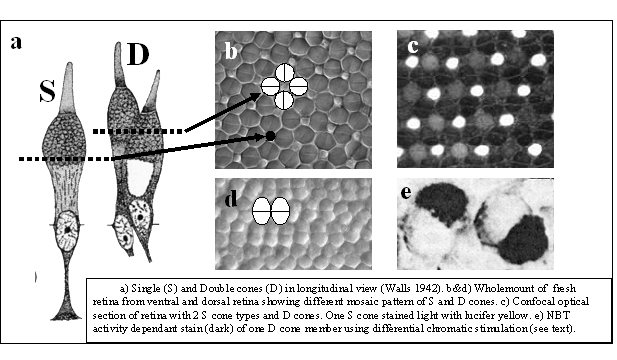
Cone photoreceptors are either single or multiple cells. Photopic systems always comprise single cones, the smallest ones being UV-sensitive, the medium ones being blue or green sensitive and the long ones being green or even red sensitive (Gouras, 1984; Bowmaker, 1991). The multiple photoreceptors are usually paired cones which occur in all vertebrate retinas with the exception of the placental mammals (Rowe, 2000). Triple or even quadruple cones are rarely described in some species and are considered to be a possible consequence of retinal regeneration (Cameron et al., 1997). The general characteristic of multiple photoreceptors is that the members share a large part of the cell membrane of the inner segments. Their outer segments are always separated and they always have two nuclei.
Paired or double cones can be either equal (percomorph fish, amphibians) or unequal (reptiles, birds, cyprinid fish) in shape, the latter consisting of a principle and an accessory member. Double cones can also be identical or unidentical with respect to their photopigments. Unequal double cones are generally unidentical. Identical equal double cones are also called twin cones (Wagner, 1990).
A short treatise on the functional significance of double cones by Mickey Rowe (2000) inspired me to write this dissertation that I dedicate to the late professor Piet Dullemeijer who never failed to point out that there are different categories of form-explanation (Dullemeijer, 1974; Dullemeijer & Zweers, 1997).
Explanation theory
It was already Aristotle who distinguished various sorts of explanations (see Aristotle’s Metaphysics, 2000). Using the classic statue as an example, he argued that its specific form is determined by i) the original sketches or its design (causa formalis), ii) the used material (causa materialis), iii) the sculptor’s skills (causa efficiens) and iv) the purpose of the sculpture (causa funcionalis). In a biological analogy these causa respectively refer to the original information leading to a specific form (c. formalis), to the structural components (c. materialis), to all factors involved in the developmental process (c. efficiens) and to the function of the structure (c. finalis).
In the original approach of Aristotle these causa are independent of each other, providing a rather static model with the ” unmoved mover” as the causa formalis. To most modern biologists, all organisms are subject to cyclic processes like ontogenetic and phylogenetic development. This generates a more dynamic model with much interaction between the causa. For instance, the design (causa formalis) is not static but adapts to environmental changes and consequently influences the other causa. Consequently, the resulting processes can better be separated into a short-term ontogenetic process (c. efficiens proximus) and a long-term evolutionary process (c. efficiens ultimus).
The most direct and simple explanation of a biological structure is a detailed description of all its features (causa materialis). The more precise the characteristics the better, without even mentioning anything about the origin or purpose of the structure.
A more elaborate explanation is either causal or teleological. Biologists use both.
The causal explanation of a structure may refer to its origin, either ontogenetic (developmental) or phylogenetic (congenial), or to the level of investigation. A reductionistic explanation of a cellular structure may involve cell orgnanells or even molecular components, as such, it would still be a more specific aspect of the causa materialis.
Research on the development of a structure most certainly necessitates the investigation of the involved molecular components like enzymes and structural proteins. Explaining the form features of a structure on basis of its ontogenetic development can be referred to as the causa efficiens proximis of the structure.
The protein synthesis via the RNA transcription and translation is embedded in the nucleotide sequences of the DNA. Using the genetic code to explain the form features of a structure can be referred to as the causa formalis of the structure (see also Vinci & Robert, 2005). The genetic code can be very fundamental within an animal class or highly variable even among close relatives. What’s more, if an altered genetic code would produce an adaptive structure (with respect to its form features or development), the causa formalis would have changed, and so would the structure. Nevertheless, small differences between structures could be reflected in their designs.
Adaptation does not only concern the entire organism but also the structure in relation to its direct environment. Therefore, the developmental change of a structure – on either time level – is best explained as a transformation that is determined by constructional constraints and functional demands, the latter referring to a functional explanation of a structure, the causa finalis. The purpose of a structure may involve physiologic, behavioural and environmental attributes that serve the survival of the organism to which the structure belongs on a short-term. Environmental changes may alter the functional demands that are made to obtain best circumstantial fitness (causa efficiens ultumus that on the long-term alters the causa formalis).
A functional explanation of a structure involves the properties of the individual structure itself. Explaining a structure in relation or in combination with its (structural) surroundings necessitates a more holistic approach and would provide a synergistic explanation as is used in for instance constructional morphology (Barel, 1983; 1993).
What sense has the separation of explanations? It seems all kind of artificial. And indeed, it is as artificial as it is to separate the synthesis of a protein from the physiological process it catalyses or to separate a piece of tissue, an organ, from an entire organism, or a population from its ecosystem. Still, such abstractions have proven to throw more and more light on an initial obscure landscape by the exploration of its subdivisions.
Most biologists are not satisfied with one of the explanations as described above. They try to combine causal and functional elements to provide an evolutional explanation that pairs gene expression and developmental clues to fitness and fecundity of the specific population that exhibits the structure under investigation. This is a time consuming procedure and it will need the skills and dedication and above all the mutual understanding of seemingly opponent disciplines and methodologies to make progress in this otherwise traditional field of animal morphology.
Explaining double cones
In understanding the double cone as a morph in comparison to the single cone – why one double instead of two singles? – one must focus on the properties reserved to the double cone. Properties that equally can be realized by single cones are insufficient contributions to a satisfying explanation. Following the assumptions of other investigators, many properties will be mentioned as they are important to the visual system and possibly not well understood even if they are not reserved to double cones. At the risk of labouring the obvious, I wish to emphasize that I have not the intention to explain the processes of photoreception if they can be performed with mere single photoreceptors.
Causa formalis by means of genetic coding
One may presume that some sort of creative power used the double cone as an essential component in its intelligent design of the retina in the majority of the vertebrate kingdom. Yet, it seems more likely that the blueprint of the double cone is embedded in a great number of codon sequences located on one or more chromosomes.
The presence of double cones in the retinae of most vertebrates, with the exception of the placental mammals, indicates a conservative genetic basis for their origin. The disentanglement of the genetic code that underlies the morphogenesis of the double cones will provide one legitimate explanation of the double cones.
Perhaps needless to say, but the genetic code is probably quite complex and certainly not (yet) available. There is, however, an indirect way to investigate the significance of double cones through congeniality. Is the presence of double cones restricted to vertebrates? So far, I haven’t read any scientific report mentioning otherwise. Ancestors of the early chordates, the agnates, have well developed eyes. The presence of double cones in the retina of the lamprey would accentuate their conservatism and their blueprint must then already be present in the genetic code of these primitive fishes. On the other hand, the agnates may lack double cones which leads to the next question: what other resemblance is there among vertebrates that lack double cones? Chondrichthyes (cartilaginous fish like sharks and rays) and placental mammals don’t possess double cones. They may have in common that they probably came into being more recently than for instance bony fishes, but certainly way before birds, that do possess double cones. Often mentioned with respect to the absence of colour vision in mammals – only primates have colour vision – is the nocturnal behaviour of the mammalian ancestors. Could this also be linked to the absence of double cones? Unfortunately, nocturnal behaviour as well as obscure habitats are described for various vertebrates that do possess double cones (owls, gekko’s, toad, pikeperch, deep-see fish). Actually, I’m anticipating a functional approach by using the causa efficiens ultumus to suggest a causa formalis.
Causa efficiens proximis by means of cellular or retinal development
The occurrence of double cones have raised the question whether they originate by incomplete fission or by partial fusion of single cones. The development could directly be linked to certain proteins [lower level] or to (parts of) neighbouring cells [similar level].
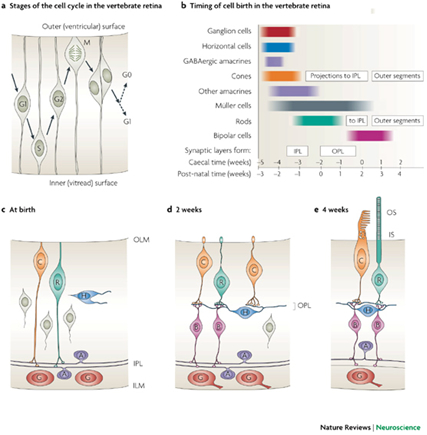
Early development of the retina starts from inside out with the ganglion cells to cones dominating rods and from centre to periphery where new neurons, except rods, are added from the retinal margin. The latter process continues throughout life in fish and some amphibians.
It cannot be excluded that the morphogenesis of the (double) cones and postsynaptic neurons along the retinal periphery requires an organizing coherence in order to develop a well functioning cup-shaped structure. The requirement may utilize the (cross-sectional) oblong shape of the double cones to realize retinal growth in a similar way as a fishing net would be unfurled. Accordingly, the double cones are always parallel along the retinal margin and often in a more rhombic and even perpendicular orientation in the stretched major surface of the retina. Whether the oblong shape of the double cones is a cause or a consequence of this process is still unclear. Besides, this cannot explain the row mosaics as found in many species.
Recent investigations on the embryonic development of vertebrate photoreceptors are still far from an identification of a so called double cone stimulant. In general, the morphogenesis of photoreceptors is linked to a thyroid hormone, to neuroD, and to other proteins (Yan & Wan, 2004; Baniahmad, 2002; Stenkamp & Cameron, 2002; Morrow et al., 1998). During ontogenetic development of the retina in various fish species, the developing rods and cones seem to play a prominent role in the determination of the neighbouring photoreceptors, resulting in a regular mosaic (Raymond & Barthel, 2004; Stenkamp et al., 1996)). The proposed synergetic mechanism, however, didn’t provide an explanation for the origin of the double cones yet.
Determination of signal proteins and the allied receptors that induce the morphogenesis of the double cones will provide another legitimate explanation of the double cones.
Causa materialis by means of structural features
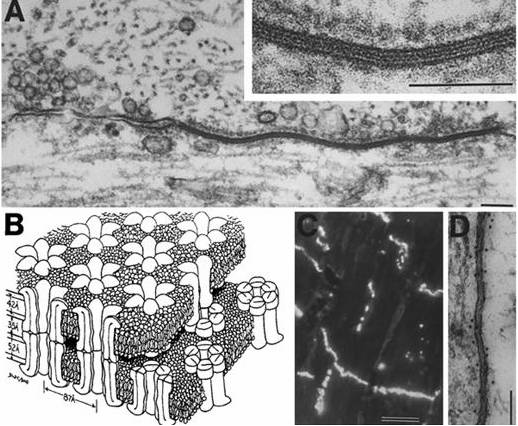
The only structure that double cones truly distinguish from single cones is the joint membrane of their inner segments. The causa materialis of the double cone should therefore be focused on this feature. The joint membrane may have its influence on the distribution of the mitochondria since most of them are concentrated in its vicinity (Rowe et al., 1994). This heterogeneous distribution of organels with a relatively high refractive index may have some influence on the way a light-beam is guided through the inner segments. Since the refractive index is the same at both side of the joint membrane, the light guidance in a double cone is unified and the two members are called to be optically coupled (Rowe et al., 1994).
As in fibre optics, the amount of light energy per unit time depends on the cross-sectional area of the photoreceptor: larger cones do have a better photon catching ability (Van der Meer & Anker, 1984) or quantum catch efficiency (Kröger et al., 1999) than smaller ones. A large single cone with equal cross-sectional area as a double cone would generally need less cell-membrane than that double cone (certainly when its an unequal double). So, what would be the benefit of a double cone over a single one? Here, another salient property of double cones comes up for discussion: most double cones, at least all unequal ones, possess two different photopigments.
Thanks to the optical coupling, the guided light beam in a double cone can be partly absorbed by two sets of different photopigments. In view of the microspectrophotometric measurements of the two members of most double cones (Bowmaker, 1991), it seems obvious that a joint venture of the two members of a double cone results in a broader spectral absorbance of a guided light beam.
Another property of the cell-membrane, and thus of the joint membrane, may be the (partial) polarization of transmitted and/or reflected light (Cameron & Pugh, 1991). If so, this may have effect on incoming light that already has some degree of polarization. How this may influence visual detection will be discussed below.
Causa finalis by means of colour vision
Double cones never contain short wave photopigments. Even in the absence of single cones their absorbance maxima are usually above 500 hm.. This does not mean, however, that double cones are necessary to carry middle- and longwave photopigments as these are carried by single cones as well, even in the presence of double cones with the same photopigments (Bowmaker, 1995).
Simultaneous use of at least two different photopigments is a necessary condition for colour vision. Di- tri- or tetrachromatic vision does not automatically imply the ability of colour discrimination. The various photopigments could just as well serve optimal contrast (contrast hypothesis; Lythgoe, 1979).
The opponent-process theory of colour vision (Jameson, 1972) involves integration of synaptic signals which can be either antagonistic (opposite synaptic effects) or synergistic (equal synaptic effects). Accordingly, integration of the inhibitory effect of the one double cone member and the excitatory effect of the other member provides antagonistic signals carrying colour information along the “green-red” opponent colour channel. Integration of synergistic signals from both double cone members is assumed to enhance contrast information along a luminance channel (for a review, see Nuboer, 1986).
So, in spite of the optical coupling of the two members of the double cones, they may also follow different neuronal pathways (Haesendonck & Missotten, 1979; 1984) and the cone-specific horizontal cells play an important role in the development colour constancy (Stell, et al., 1994; Verwey et al., 1996; Kamermans et al., 1998). So, it seems plausible that the double cones attribute to colour vision in many vertebrates when enough light is available. At least in fish, behavioural experiments give further evidence assumption (Siebeck et al., 2008; Neumeyer, 1992). It remains, however, unclear why this function could not be performed by mere single cones as in primates.
Causa finalis by means of improved resolution and movement detection
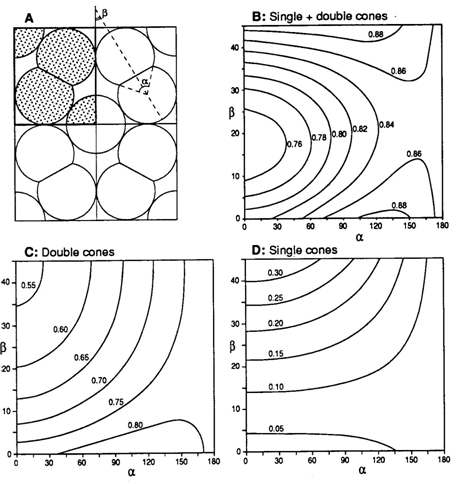
Visual resolution is the ability to distinguish two spots from one another which is done by at least two photoreceptors separated by a third one, provided that each of these photoreceptors has its own visual pathway (Johnson, 1960). According to this condition, resolution will be improved by an increase of cone density. If the two members of a double cone were to function separately this could be applied to them as they are more closely packed than if they were separate cones. Since the double cones constitute the lattice of the cone mosaic, they appear to serve maximization of the cone packing (Van der Meer, 1992). Especially in combination with single cones, but also in pure double cone mosaics, the packing is improved in comparison with a corresponding cone area realized by circular elements only.
These considerations only hold if the members of double cones follow different pathway (see above), which they may do under specific (bright) circumstances.
However, if the double cones are optically coupled, the opposite is the case as a double cone takes more room and the cone-density, as a consequence, decreases. For the same reason, movement detection as a function of resolution can neither be improved by mere double cones in spite of what some authors may have believed (Lyall, 1957; Wagner, 1978; Campenhausen & Kirschfeld, 1998; Schwab, 2002).
Causa finalis by means of increased sensitivity
If the members of a double cone are optically coupled, the double cone can be regarded as a functional unit and the signals from both members are added and follow the same pathway for the benefit of sensitivity. The hypothesis is supported by the electrical coupling between the members of double cones (Richter & Simon, 1974; Miller & Korenbrot, 1993) although this may not be true in every species (Attwell et al., 1984; Lasater et al., 1989). Electrical coupling doesn’t seem to be restricted to the double cones and can simply be realized by gap junctions between all adjacent photoreceptors (Zhang & Wu, 2004). On the other hand, experimental work on isolated double cones containing different visual pigments (533 nm and 620 nm) “suggests a prominent role of the double cones in luminosity rather than wavelength detection” (Marchiafava, 1985).
Visual sensitivity can be understood in various ways. Here it is meant as the threshold determined by the minimum photon catch necessary to generate a neuronal pulse. The larger a photoreceptor the higher its photon catching ability and the lower its threshold, provided the same photopigment and visual pathway. According to this condition, the optically coupled members of a double cone are about twice as sensitive than if the members were separated single cones. On the other hand, if the two separate single cones would follow a joint pathway, there would practically be no difference with a double cone. Yet, due to their cellular properties, single cones can never reach the size that double cones can. The connected large photon catching ability would certainly be an advantage in an obscure, e.g. underwater, environment. Large double cones are a common feature in benthic fish and nocturnal amphibians.
Next to this capacity to enlarge the photon catch, double cones have yet another way to lower their the threshold. Most double cones possess two different photopigments. Since a double cone can be apprehended as an functional unity following a luminance channel (Devalois, 1973; Marchiafava, 1985; Nuboer, 1986), they have become photoreceptors with a broader spectral sensitivity than any single one. Photons are characterized by their wavelength and the molecular composition of a photopigment determines which wavelength is maximally absorbed. The maximum absorbance of a photopigment is always strongly marked (Bowmaker, 1991) so the possession of two different photopigments could certainly be an advantage. In equal double cones this advantage may be slight, in unequal double cone it could certainly make a difference. Many photons passing the photopigment of the smaller member will be absorbed by the photopigment of the larger member of one light guiding system. The slender double cones in e.g. diurnal birds may in this way serve a better demand of photon catch without loss of resolution.
Causa finalis by means of magnetic orientation
Many behavioural studies have established that birds are sensitive to the global magnetic field and its variations (Wiltschko & Wiltschko, 1995; 2003). In a variety of vertebrates, mostly birds, specific wavelengths of light have been shown to influence magnetic orientation. The avian magnetic compass receptor appears to be a light-dependent, wavelength-sensitive system that functions as a polarity compass (i.e., it distinguishes poleward from equatorward rather than north from south) and is relatively insensitive to changes in magnetic field intensity. The receptor is within the retina and is based on one or more photopigments, perhaps cryptochromes (Beason, 2005). Beason & Swali (2001) proposed that double cones could act as magnetic receptors. Exactly how the double cone would be able to function as such remained unclear but the wavelength sensitivities of investigated bird species corresponded to the sensitivities of the two members of the double cones. Recently, an integrated model has been proposed that combines the colour gradient compass and the magnetic compass (Kragh Jensen, 2010). In the model, the large range of colour gradient (370-610 nm) could very well be smaller, especially when polarisation sensitivity is included.
Some explanations relate to the mosaic rather than to separate double cones. Such synergistic approaches are also considered as functional explanations.
Causa finalis by means of polarization detection
Since double cones may show a perpendicular position in the cone mosaic, they were repeatedly appointed as photoreceptors for various directions of polarization (Cameron & Pugh, 1991; Rowe et al., 1994; Hawryshyn, 2000).The idea came from the structural feature that the mitochondria of a double cone are concentrated next to the joint membrane between the two members, allowing them to act as a single waveguide with an elliptical cross section. This may cause the double cone structure to act as a birefringent receiver and a linear polarizer, allowing vertebrates with double cones to separate orthogonal linear polarizations of light and subsequently substract these signals to output a polarization-difference image (PDI). Although a biological hypothesis, it has certainly inspired technicians to apply PDI in cameras and target detectors (Rowe et al., 1995; Bains, 1996). The technique has improved the extraction of images under veiling light due to scatter.
Many vertebrates, under water or in the air, have to navigate through densely scattering media and would certainly profit from PDI to enhance their target detection. Some investigators suggested that polarization detection is generated by the joint membrane of the double cone that would act as a dielectric mirror and that polarization dependent reflection and refraction at this membrane constitutes the underlying mechanism in the transduction of polarization into intensity variation at the photoreceptors outer segments (Novales Flamarique et al., 1998). So far, studies on fish species with a square pattern show polarization sensitivity whereas species with a row pattern do not (Novales Flamarique & Hawryshyn, 1998; Degner & Hawryshyn, 2001)
Polarisation detection hardly exists in humans and there are no reports of its presence in other mammals lacking double cones. Birds, especially pigeons, are considered to be able to discriminate the field of polarization, mainly due to the work of Delius (Delius et al., 1976), which, however, has been criticized and may not be entirely reliable (Coemans et al., 1990).
Behavioural experiments on sunfish indicate that they cannot be trained to discriminate light on the basis of the polarization state even though the same fish in the same apparatus can be trained to make colour discriminations (Rowe, 2000).
Causa finalis by means of optimised cone packing and transformation capacity
The cross-section through the inner segment of a double cone, acting as a wave guide, resembles two overlapping circles, varying from almost circular to the elongated eight-shape. In combination with circular single cones, equal double cones may provide a better occupation of the retinal surface (depending on the amount of cross-sectional overlap) thus optimising the photon catch of the photopic system (Van der Meer, 1992). The two most common regular mosaics are the so-called square-pattern (single cones surrounded by perpendicular arranged double cones) and the row-pattern (parallel orientated double cones). Transformation from a tri-chromatic square-pattern into a row pattern of mere large double cones favours the photon catch at a limiting spectral bandwidth, which suggests the mosaic to function as a wave collector of an either broad or narrow spectrum. In some fish species that, as part of their life cycle, migrate to deeper water, such a transformation has been observed (Boehlert, 1978). Such a transformation with optimal photon catch is impossible with mere circular single cones (in cross-section).
Many irregular mosaics consist of a number of different single cones and unequal double cones. In cross-section their inner segments may all be approximately circular shaped. Retinal cross-sections, however, reveal that the inner segments of these cones vary in length and/or position (e.g. Kunz, 2006). Short, medium and long sized single cones are combined with the principle and accessory members of unequal double cones in retinal rows with high packing and stratification of photopigments (Bowmaker, 1995). In this way the packing seems so high that there is hardly any room left to allow rod myoids to pass. It has been suggested that the stratification of photopigments undo the chromatic aberration of the optics. This, however, would contradict the wave guide theory which presumes an image projected at the limiting membrane. Besides, not all stratifications consist of similar photopigments.
It is questionable, whether, in this context, the major function of the unequal double cone is to maximize the packing, as such a function could be performed by mere single cones as well.
Causa efficiens ultumus by means of naturally selected functions
When double cones perform a function that benefits the fitness of the individual organism, natural selection is likely to spread the function – and thus the coherent double cones – among the population that the organism belongs to. This may have happened among populations of completely different species under completely different circumstances. It is, indeed, quite possible that more than one function has developed. To birds of prey it may have been primarily an increase of photopic sensitivity without loss of acuity, to salmon it was perhaps magnetic orientation to support their migration movements. To juvenile fish it may have been the maximization of the photoreceptor packing in order to improve their visual abilities, to adult fish it was perhaps the development of polarization sensitivity to observe through a scattered medium. To some non-mammalian vertebrates occupying a bright environment, double cones may have primarily been useful as opponent colour detectors, whereas to other species, migration towards a less colourful habitat was supported by the transforming capacity of the double cone mosaic.
So, whatever function is focussed on, it doesn’t necessarily belong to every organism with double cones. However, what cán be expected to be similar in all these organisms, is the genetic coding and ontogenetic processes that leads to the morphogenesis of double cones.
Conclusive remarks
The genetic code that underlies the morphogenesis of the double cones is still completely out of sight. Necessary DNA-sequences for the benefit of the synthesis of enzymes that are involved in this morphogensis are not known, neither is there a notion of much interest in that direction. After all, humans have no double cones, much less related affections.
As mentioned earlier, the various causa are linked together: functional explanations depend on structural features and they are considered functional as the related properties are supposed to serve fitness and fecundity of a population. The properties must, therefore, be inheritable which means that the genetic code provides the necessary proteins to realize the structural features that are used in the functional explanations.
And indeed, there is more than one functional explanation. And most of them seem quite plausible too, which raises several questions. Has this multi-functionality existed from the beginning of vertebrate development and does it occur within one organism during ontogeny or even simultaneously? And would all the profits be merely accidental?
Well, coincidental advantage, isn’t that what evolution is all about?
The present information suggests, that most conclusive force comes from the double cone as a functional unit. Even though they often contain different photopigments, colour vision doesn’t seem to be their primal function. By the seat of my pants I would go for their contribution to an increased photon catch (lowering the photopic threshold), which doesn’t exclude any other extra functional significance. But this is not a proper scientific approach. Much better to await the results of the current research of the Sensory Neurobiology Group of the University of Queensland (Australia) on the question ‘What do double cones do’? (http://ilc00f.facbacs.uq.edu.au/VTHRC/ecovis/Images/double%20cones
/ARCDiscovery2004f_Esection.pdf)
< click here to go back to PAN-OPTICS & EVOLUTION |
|
|