| |
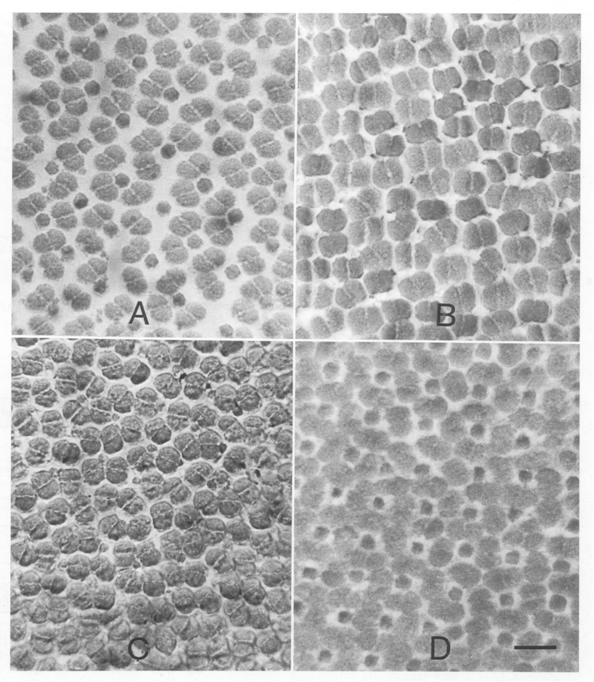
Regular geometric patterns of retinal receptors are a common feature of many teleosts (Ali & Anctil, 1976). These patterns are tessellations of morphologically different cone types and are basically characterized by the mutual orientation of the double cones (cited from Van der Meer, 1992). A limited number of tessellations appears to be highly successful considering their dominant distribution among divergent taxa: row and square patterns of the percomorph type, viz. parallel and perpendicular orientation of double cones with equally sized members. Also the intermediate rhombic pattern is often observed. Apart from phylogenetic variation, the occurrence of either one of these configurations depends on the intra-retinal position, developmental stage and/or environmental condition (Lyall, 1957; Engström, 1963; Ahlbert, 1969, 1975; Boehlert, 1978; Van der Meer et al., 1995; Fritsches et al., 2000; Reckel et al., 2002, 2003).
Several functional explanations of the receptor mosaics have been suggested varying from improvement of visual acuity and movement detection to a uniform distribution of colour specific sensitivity and polarization sensitivity (Munz & McFarland, 1977; Wagner, 1978; Ali & Klyne, 1985; Levine, 1985; Hawryshyn, 2000; Reckel et al., 2001; Fritsches et al., 2003). These proposals were primarily based on correlations between the histological data and a specific behaviour or environment rather than being derived from the optic demands due to environmental conditions. Besides, the significance of mosaic-transformation during ontogeny, which occurs in at least a number of species (Ali & Hanyu, 1963; Boehlert, 1979; Novales Flamarique, 2001), may per se be a co-determinant of the mosaic. Van der Meer (1992), therefore, used a different approach and choose to investigate the mosaics on constructional constraints and transformational demands. Accordingly, a geometrical model was derived from functional demands made upon the photoreception in fish, to predict several cone-configurations providing the best possible solution for visual perception under different environmental light conditions.
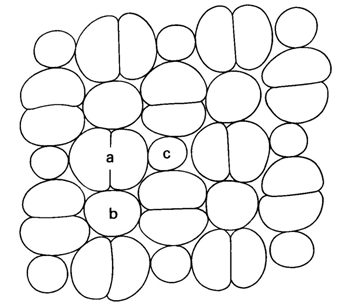
In many teleosts the cone mosaic is of the percomorphal type: basically an arrangement of four equal double cones surrounding a single cone. Referring to their location within the mosaic, the single cones are called central or accessory and either of them can be present/absent. The single cones usually contain short wave visual pigments while the double cones hold the medium and long wave pigments (Levine & MacNichol, 1979, Bowmaker, 1995). The space between the cones is occupied by the myoids of the rods connecting their sclerad inner and outer segments with their vitread nuclei in the outer nuclear layer. Also, Müller cells (radial fibres with a metabolic function) and the protrusions of the pigment epithelial cells penetrate between the cones.
Functional demands and constructional constraints
The considered functional demands made upon the photopic system were the ability to discriminate details and colours at the relatively low light levels of an aquatic environment. Detail discrimination is determined by the resolution which is considered to be a function of cone density and eye-size within the optical limits of the eye lens (Otten, 1981; Van der Meer et al., 1995). Apart from the distance between adjacent receptors, detail discrimination is assumed to be improved by colour discrimination which demands receptors containing photopigments with different absorption maxima (Loew & Lythgoe, 1978). The ability to function under dim light conditions was determined by the sensitivity of the photopic system which was considered to be a function of both cone convergence and cone size (independent of eye-size; Van der Meer & Anker, 1984)
Because cone density and cone size are form features competing for space in the plane of the receptor layer, their functions (resolution and sensitivity) are balanced and cannot be maximized simultaneously. By changing the density/size ratio of the cones the resolution/sensitivity balance can vary to meet the requirements of the specific habitat and niche of the organism, without affecting the cone packing (i.e. the fraction of retinal surface occupied by the cones). To maximize the photon catching ability of the photopic system, the cone packing should be maximized (Barel, 1993)
Since a receptor cell contains only one type of photopigment, colour vision demands physiologically different cones (Lythgoe, 1979). Consequently, the retinal area is divided among two or more different colour-specific cone types at the cost of both the mutual distance between similar cone types (and thus the colour-specific resolution) and the absolute sensitivity of the original system (photopic threshold, Van der Meer, 1994). These colour-specific cone types are also competing for space. Loew & Lythgoe (1978) pointed out that two different visual pigments (one matching the background light and one matching the light reflected by the object to be detected) provide an essential improvement. The addition of other photopigments may improve colour discrimination and perhaps also detail discrimination, whereas the restriction to only one photopigment may improve the absolute sensitivity within a more limited spectral domain.
The ratios between cones and ganglion cells and the connectivity patterns (Wagner, 1990; Mack, 2006) suggest the existence of photopic units consisting of a limited number of cones converging to one ganglion cell. For the benefit of resolution the number of comprising cones should be small.
The model
As a start, in a wide-angle range of vision movement detection in all directions the visible field is considered to be important. Since movement detection depends on the distance between the contiguous units, the distances between the centres of neighbouring units should be as constant as [possible to provide a high degree of homogeneous resolution (DHR). This requires, together with the demand to maximize the packing, an arrangement of photopic units according to a lattice that provides a tessellation of congruent regular polygons. Rectangular or rhomboidal polygons are accepted as well, having still equal distances to four of their eight neighbours along two different lines. The order of symmetry of the resulting pattern unit is a measure to indicate how well this requirement (homogeneous resolution)is met.
Detail discrimination can be improved by colour discrimination, provided that the latter is performed on the same level of resolution, i.e. a photopic unit should include the different colour specific cones that are present in a specific retinal area. The abovementioned arguments with respect to movement detection are equally relevant to the separate colour specific cone types. Therefore, a homogeneous distribution of separate colour specific cone types (HDC) is essential. This requirement is met when the symmetry of the unit holds also for the physiological differences (in the mammalian eye, where double cones are missing, for that matter, a random distribution of colour specific cones would provide the best solution; Li & DeVries, 2004; Mack, 2006).
Since the spectral distribution in an aquatic environment depends on variables such as depth, suspended and dissolved matter, scatter, etc., the colour sensitivity of the unit should be able to cope with such a variation. Therefore, a high variation of the area ratio of the different cone types within a unit would be advantageous. This is simply performed by changing the area ratio (and thus the sensitivity ratio) of the single and double cone types within a unit. For an optimal design the range of this colour-sensitivity balance should be maximized with minimal variation of the packing.
Light projected on the retina could easily be scattered from one cone to its neighbours. At the level of the outer segments the reduced contrast would negatively effect the resolution. The cones, especially their outer segments, should be separated by means of a light-absorbing substance (melanin pigment within the protrusions of the pigment epithelial cells penetrating between the cones). Intercellular space is also necessary for metabolic reasons (Müller-fibres). Last but not least, the opposed migrating movements of rods and cones require separation of the cells while some space between the cone remains necessarily occupied by the rod myoids (especially in the larger eyes of adults since new rods are generated throughout life between the other retinal neurons; Johns & Fernald, 1981). Since separate oblong cells are likely to have curved shapes (in cross-section), their sections in the plane of the receptor field are assumed to be basically circular. The minimal space between touching circles is regarded to provide sufficient intercellular space.
In summary, the following conditions, premises and functional demands were used in producing the model:
- separations of elements entails a curved shape (condition)
- small, uniform and symmetrical photopic units (condition)
- two different colour specific cone types (premise).
The demands are met by
- minimizing the size of the units (high resolution)
- maximizing the symmetry of the unit (improves movement detection)
- maximizing packing (high photon catching ability)
- maximizing variability of the cone’s area ratio (potential transformation)
It should be noted that compromises between the demands and the premise can be realized and that several solutions are likely to occur.
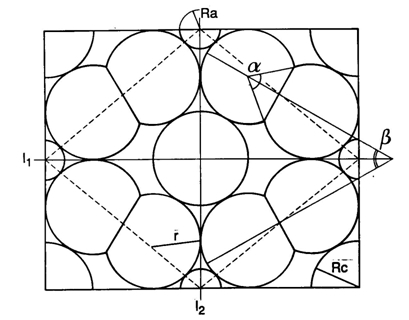
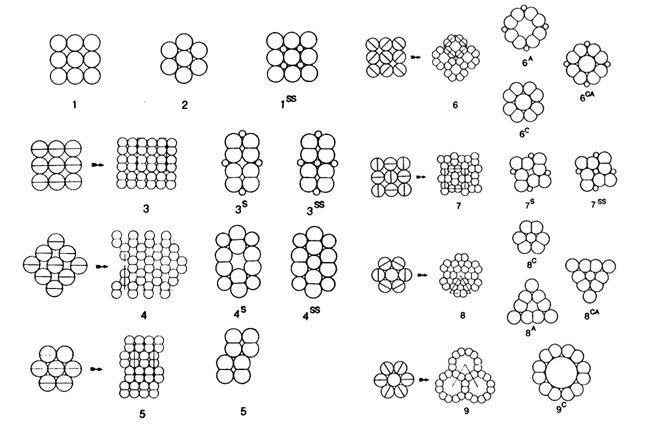
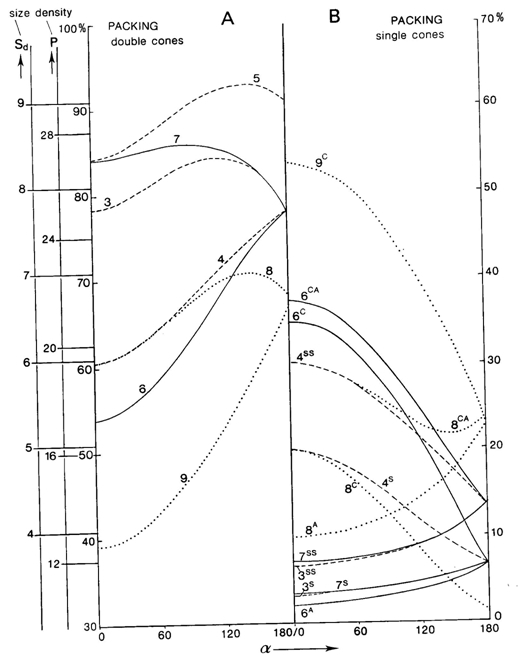
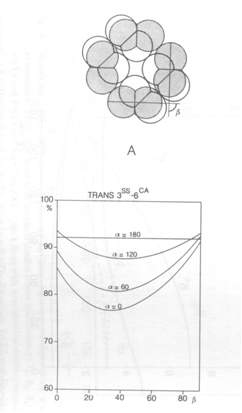
According to the mentioned conditions, premises and demands, a number of mosaics were studied, varying from mere single cone patterns, combinations of single and double cone patterns to mere double cone mosaics. The model depends on only two parameters (cited from Barel, 1993): alpha for the shape of the double cones and bèta for their mutual orientation and hence for the configuration of the mosaic.
Computations show that a double cone with alpha = 115 degrees provide the best possible packing of 84%. In comparison, a row arrangement of single cones (alpha = 0 degrees or 180 degrees) provides a packing of 78,5%. A triangular arrangement of single cones could provide a better packing (91%), but this arrangement does not meet a number of demands set by the model.
Combinations of double and single cones provide the best packing (94%) in a non-existing square arrangement with small single and additional single cones (alpha = 112 degrees). The known square pattern of four double cones surrounding a central single cone has a highest packing of 90% (alpha = 83 degrees). With accessory single cones as found in juveniles of some other species (Lyall, 1957) the maximum packing comes to 93% (alpha = 107 degrees). Moreover, these mosaics can simply be rearranged into a row-configuration by changing the mutual orientation of the double cones (bèta) over 45 degrees. The resulting maximum packing would remain equal (93% at 139 degrees). Even if alpha was kept constant at for instance 120 degrees, this would not effect the packing of 93%. However, in the row-configurations as found in cichlids, the single cones are usually missing, providing a maximum packing of 84%. By a slight rearrangement of the rows, the packing could rise to 93% again, which is probably never realized as row mosaics were always found in adult benthic species with high numbers of rods that need the intermediate space between the cones.
Transformation of cone patterns were repeatedly reported and can be understood as an adaptation to a changing photic environment as the fish migrates towards deeper water. The small accessory UV-sensitive single cones gradually change their sensitivity and disappear as the fish grow older (Bowmaker & Kunz, 1987; Cheng & Novales Flamarique, 2007). Square mosaics with large blue sensitive single cones may transform into entirely red and/or green sensitive row mosaics as the fish occupies a photic habitat where the medium and longer wavelengths become more dominant (Boehlert, 1979; Kusmic & Gualtieri, 1999; Witte, et al., 2005).
In a few species the rather rare triangular mosaic was observed. The assumption that this pattern is especially fitted to detect small movements as a result of high resolution and high symmetry – provided that the double cones are identical, which they are indeed (Levine & MacNichol, 1979) – is supported by their occurrence in the retinas of the Esoxidae, a family of specialized ambush-hunters (Wagner, 1972). The triangular pattern was also mentioned to occur in the retina of the rudd Scardinius erythrophthalmus (Scholes, 1976; Nag & Bhattacharjee, 2004). However, the presented model is restricted to the percomorph retina (as in cichlids) and takes no account of unequal double cones as found in Ciprinidae and various other fish families.
|
|
|