| |
The retinal form variations among haplochromine cichlids could be the result of phenotypic plasticity: environmentally induced acceleration or retardation of development of individual parts relative to other parts without alteration of the genetic program of development (Smith-Gill, 1983). The phenotype can vary within the limits set by the genetically determined ‘norm of reaction’, which could be a significant mechanism in the evolutionary development (Schlichting, 1986; Stearns, 1989). In more general terms phenotypic plasticity is the production of multiple phenotypes from a single genotype, depending on environmental conditions (Miner et al., 2005). A wide diversity of organisms express phenotypic plasticity in response to biotic and abiotic aspects of their environments (DeWitt & Scheiner, 2004).
It is a common feature in cichlids (see Witte et al., 1990 for a review) and several studies of the visual system in cichlids have demonstrated the occurrence of light-induced phenotypic modulation (Zeutzius & Rahmann, 1984; Zeutzius et al., 1984; Van der Meer & Anker, 1986; Kröger et al., 1999, 2001, 2003).
Haplochromine cichlids differing in their vertical distribution (Goldschmidt et al., 1990) have to cope with light conditions that differ both in intensity and in spectral composition (cited from Van der Meer, 1993). The retinas of these species reveal variations of form-features such as size, number and arrangement of the photoreceptors, which may be visual adaptations to the photic conditions of their species-specific habitat (Van der Meer, 1989).
Phenotypic plasiticity of the visual system in cichlids may concern at least two environmental factors, viz. light intensity and spectral composition. Although a long period of absolute darkness is hardly a natural condition in the studied fish, light deprivation may provide one extreme of the phenotype, while colour deprivation may add relevant information concerning the effect on the different colour-sensitive photoreceptors.
Procedure
Mouth-brooding females of H. sauvagei were placed in light-proof tanks (ca. 55 l; 25oC + 1oC) and removed after three weeks when their fry were “free-swimming”. One group of larvae was raised in complete darkness (D-spec.), one group was raised in blue light (B-spec.), one group was raised in red light (R-spec.) ad one group was raised under usual laboratory conditions (ca. 150 lx; L-spec.). In the case of the R- and B-spec., the light source consisted of one Philips TLD 18W/33 suspended 5 cm above a diffuse glass pane covered with one Lee filter (26 x 51 cm), no. 106 (red) and no. 119 (blue), respectively. The filters were placed 25 cm above the water surface, providing an illuminance of ca. 1 – 5 lx in the tank. The fish exposed to light (L-, B- and R-spec.) had a diurnal light/ dark rhythm.
Periodically, a few specimens from each group were exposed to normal room light for half an hour (to make sure that all the retinas were equally light adapted) and treated according to the histological procedures described elsewhere (Van der Meer, 1993).
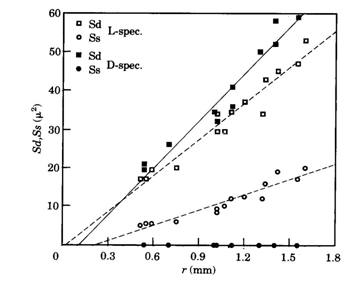
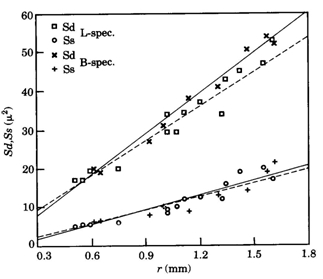
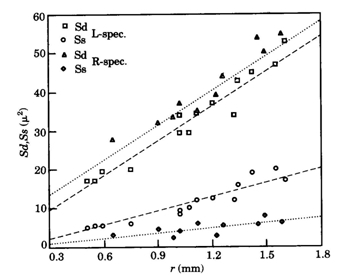
The size of either a double cone (Sd) or a single cone (Ss) was derived from the tangential sections and calculated from the elliptic and circular shape of the cross section through its inner segment. The area of a photopic unit (Ac) comprising two double cones and one single cone (Otten, 1981) then equals 2Sd + Ss.
The angular cell density (H; number per unit visual angle) was calculated using:
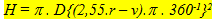
where v is only relevant in calculating the angular ganglion cell densities (Hg) of small eyes.
To compare retinal development under different light conditions, multiple regression analysis was applied using dummy variables to indicate the different light conditions (Kleinbaum & Kupper, 1978). Two regression equations of the form y = Bo + Bx+ E were combined to form the multiple regression model:
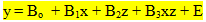
where z represents the condition. The F-values were used t test for the significance of parallelism and coincidence of the two original regression lines:
hypo: B3 = 0 (parallelism):

hypo: B2 = B3 = 0 (coincidence):

These F-values were compared with the statistical table of Fi,j,0,95, where I is the degree of freedom of the regression (1 or 2), j is the degree of freedom of the error o the model (depending on the number of data) and 0,95 is the confidence limit.
Light deprivation
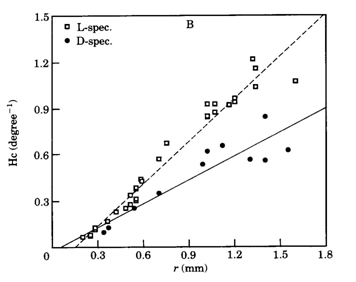
The rearing of cichlids in complete darkness has differing, probably species-depending effects on their visual system. Grün (1979; 1980) observed a light induced acceleration of early retinal development in Tilapia leucosticta which led to the retardation of postsynaptic neurons like ganglion cells in light deprived specimens. A similar effect was found in the tilapia Sarotherodon mossambicus (Van der Meer & Anker, 1986) which species also showed severe impairment of visual acuity together with structural changes of the optic tectum (Zeutzius & Rahmann, 1984; Zeutzius et al., 1984). Van der Meer & Anker (1986) also measured an enlargement of 17,5% of the pupil size in S. mossambicus raised in darkness for a period of twenty weeks.
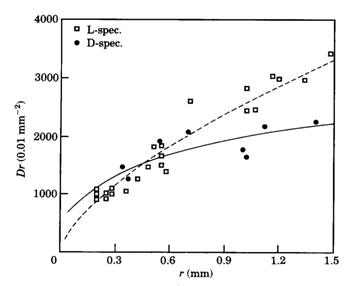
In contrast to these findings, the postsynaptic cell densities in light deprived specimens of Haplochromis sauvagei were not significantly different from those in normally raised specimens. However, the cone density remained lower than in the L-specimens. Complete absence of the single cones only partially accounted for this since the angular density of merely double cones was still significantly lower than in the L-specimens. Opposite to the severe negative influence on the single cone size, light deprivation had a positive effect on the size of the double cones, which were enlarged by a factor of 1,12 in juveniles to 1,23 in adults. The decreased area occupation consequent on the single cone reduction in the D-specimens was only partly used by the enlarged double cones through a rearrangement of the mosaic. The mean cone packing dropped from 72% in L-specimens to 60% in D-specimens. The extra available space was not occupied by an increasing number of rods. On the contrary, the rod density was also significantly lower in D-specimens compared to L-specimens.
Such a reduction of rod densities in adult stages has also been observed in light deprived goldfish Carassius auratus (Raymond et al., 1988) which these authors attribute to a degeneration of already differentiated rods.
The contrasting results of the tilapias and H. sauvagei demonstrates how careful one should be in extrapolating the effects of environmental induction, even among species from the same family.
Colour deprivation
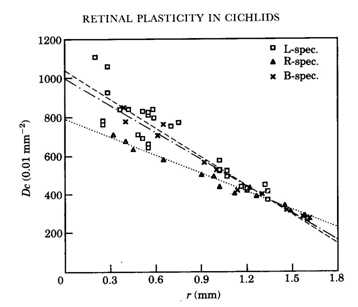
In H. sauvagei only deprivation of the shorter wavelengths appeared to affect the retinal morphology. In adult specimens, raised in a red light regime, the double cones became slightly larger than in normally raised specimens, while the single cones were considerably smaller than in L-specimens. In the central regions, the cone mosaic had rearranged from square to row and throughout the retina the cone packing had dropped to an intermediate value (69%) between the packing in the L- and D-specimens.
The photopigments were not expected to change under the influence of colour deprivation since microspectrophotometry in various species that were raised for several generations under bright laboratory conditions showed distinctive differences among these pigments (Van der Meer & Bowmaker, 1995; Carleton et al., 2005). This assumption was affirmed by studies on Aequidens pulcher after rearing in various monochromatic light regimes (Kröger et al., 1999). Exposure of this species to the different monochromatic lights appeared “to have little or no effect on the lmax of the various spectrally distinct photopigments”. In contrast, however, to colour deprivation in H. sauvagei, it is the absence of the longer wavelengths that caused a reduction of single cones in A. pulcher. When raised in monochromatic blue light “the outer segments of the double cones were longer” and “nearly 20% of the single cones were missing in the central retina”. Kröger et al. explained these results as a retinal respond to spectral deprivation “in a compensatory fashion aimed at balancing the input from the different cone types to second order neurons”. Once again, the contrasting results between H. sauvagei and A. pulcher warns us against hasty conclusions.
The reduction of single cone area in blue deprived haplochromines can be explained as a functional response since the central single cones usually hold the short-wave sensitive photopigments (Bowmaker, 1990). The observed decreasing difference in cone density during growth of the R-specimens compared to the L-specimens suggests that the initial retardation of cone genesis is later recovered through accelerated mitosis. The enlargement of the double cones could be a secondary effect due to the increased intercellular space provided by the reduced single cone area. Pressing together of the cones during their accelerated genesis could be the mechanism to the transformation of square to row mosaic. From a functional point of view, the reduction of short-wave sensitive single cones in the absence of blue light would favour the medium/long-wave sensitive double cones. A causal explanation of the short-wave light-induced modulation is not at hand.
If there is a continuous relationship between the ambient brightness and the photoreceptive area, the influence of a very dim environment would only in a quantitative sense differ from the effect of complete darkness, which seems to be maladaptive in view of the lower cone packing and rod density. The double cones, however, may play a prominent role in luminosity rather than wavelength detection (Marchiafava, 1985). In that respect, the reduction of single cones in favour of the enlargement of the double cones can be conceived of as adaptive.
The light induced morphometric changes in the retina of H. sauvagei resemble the retinal variation as observed in other haplochromine cichlids from various photic environments (Van der Meer, 1989). The species living in deeper water have tiny or no single cones and enlarged double cones and even their rod density is often lower than in a surface-dwelling species. It does not necessarily mean that these interspecific differences are the result of phenotypic plasticity. Certainly not, if the rather moderate level of plasticity in H. sauvagei is considered in comparison to the interspecific differences. If, nevertheless, the light-induced phenotypic modulation in H. sauvagei can be extrapolated to other haplochromine cichlid species, the reaction norm may have evolutionary significance (Price at al., 2003). As the fitness of specimens in the periphery of their habitat is improved through adaptive plasticity, there may be a growing feasibility for peripatric speciation, e.g. by mutation of the reaction norm (Van der Meer, 1993). Single cone reduction for the benefit of double cones may improve both luminance and red/green sensitivity which, in the natural environment of Lake Victoria, is exactly what is needed for those species living in the deeper regions of the lake as well as in the more turbid water due to eutrophication.
|
|
|