| |
Spectral preference and adaptation
Visual pigments
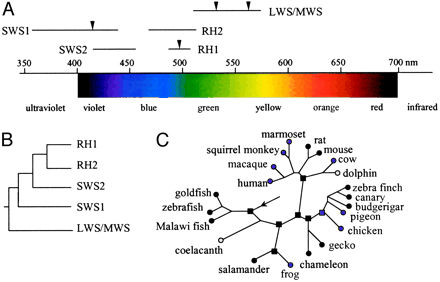
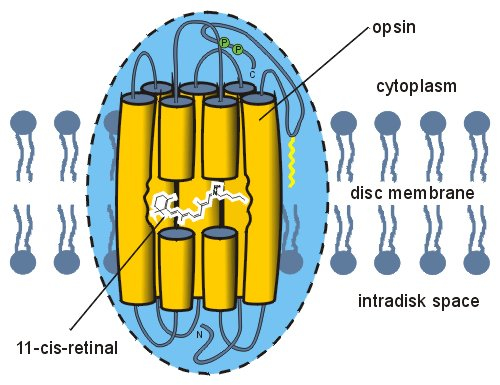
Half a billion years of vertebrate evolution have produced only a few spectrally distinct classes of cone pigment encoded by distinct opsin genes (Bowmaker & Hunt, 2006) with absorption maxima varying between 350 nm and 570 nm. This suggests that the existing photopigments are most successful. They are composed of a protein moiety, opsin, linked to a chromophore, retinol, a derivative of vitamin A. The spectral sensitivity class of the pigment is determined primarily by the interactions of retinol with specific amino acids in the protein. Invertebrate visual pigments are basically constructed in a similar way and only differ slightly in detail from the vertebrate ones (Nakagawa et al., 1999).
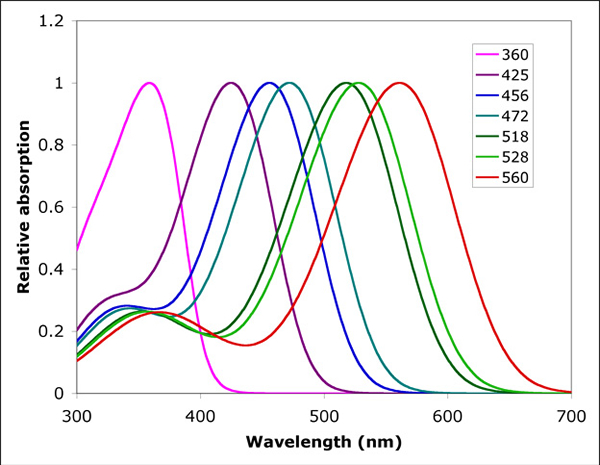
Light perception depends on the spectral bandwidth of the visual pigments. A visual pigment has an absorption maximum at a specific wavelength and a spectral bandwidth with 50% absorbance over roughly 110 nm (slightly increasing with wavelength; Bowmaker & Dartnall, 1980). More efficient light absorption would involve pigments with a broader spectral bandwidth. Which were apparently not available during vertebrate evolution. Therefore, a combination of different pigments did together not only broaden the spectral bandwidth (to some this may have led to the origin of the double cone*; see also elsewhere on this site), it made colour vision possible as well.
Visual signalling
Spectral signalling is assumed to be a common feature in animals with colour vision. The animal’s spectral sensitivity, however, often deviates from our own, and even may involve wavelengths for which we are insensitive (like the detection of UV-light). Consequently, the significance of a colour pattern is not always immediately obvious, even though the colours are presumed to play a prominent role in species recognition, sexual selection and evolution (Endler et al., 2005; Brunton, 1998). Furthermore, colour vision probably contributes to visual tasks like predator avoidance and food detection, the latter especially in visually oriented foragers. Related ‘approaching’ and ‘avoiding’ colour signals may have evolved into colour preferences (Humphrey, 1976).
Colour preference
Obviously, colour preference can only exist in an animal with colour vision, i.e. an animal with the ability to discriminate between spectral wavelengths through different, simultaneously active visual pigments. Being a psychological phenomenon, colour preference should not be confused with colour sensitivity. Colour preference must have a physiological (neuronal / hormonal) basis. This property can be either innate or learned by association; in both cases the neural basis is genetically determined. Accordingly, colour preference can become a hereditary feature. There is much diversity of innate colour preference. Many naive animals prefer short wave light (e.g. UV in most flying insects, Gao et al., 2008; violet in bees, Raine & Chittka, 2007; blue in humans, Crozier, 1999; and apes, Wells et al., 2008). Others prefer longer wavelengths (yellow and green in Colorado beetles, Otálora-Luna & Dickens, 2010; orange in some butterflies, Pohl et al., 2011; and even red in some birds, Schmidt & Schaefer, 2004 and juvenile salmon, Clarke & Sutterlin, 1985). Still, all these animals seem to be able to adapt their colour preference in a profitable sense as well as to avoid unpleasant experiences (e.g. Spence & Smith, 2008; Ham et al., 2006; Clarke & Sutterlin, 1985).
Investigating colour preference in humans has yielded some interesting theories connecting physiology to ecology. The cone-contrast model is based on an evolutionary selected cone-opponent neural response in the human visual system, revealing a general preference for short wave light as well as a gender difference at longer wavelengths (females like redder colours; Hurlbert & Ling, 2007). The ecological valence theory combines evolutionary and learned components which results in a strong correlation between colour preference and association with agreeable objects (Palmer & Schloss, 2010).
Spectral sensitivity and (aquatic) adaptation
The spectral sensitivity of an animal is usually adapted to its photic environment. Such adaptations can be the result of two mutually independent tuning mechanisms. One involves (heritable) mutations of the protein, the other involves a (phenotypic) change in the compound-balance of vitamin A. Mutations within an opsin gene can alter the interaction between the retinol and opsin, leading to a spectral shift of the pigment. The spectral shift can vary from a few nanometres to dozens of nanometres, depending on the sort of mutation.
The second tuning mechanism is realized by a change in the chromophore. Aquatic animals have to deal with a highly variable light regime. In many fish species, the chromophore consists of a combination of retinol (vitamin A1) and dehydroretinol (vitamin A2). Changing the balance between retinol and dehydroretinol causes a change of the absorption maximum of the photopigment (Temple et al., 2006). A shift from A1 to A2 has the effect of shifting the lmax of the visual pigment to a longer wavelength, i.e. the animal becomes better adapted to a more reddish environment. The shift can be induced by changing levels of the thyroid hormone (Glaschke, 2011; Allison, 2004) while the influence of seasonal cycles should not be underestimated (Temple et al., 2008).
In fish oxygenated carotenoids are presumed to be utilized for pigmentation and in juveniles the dietary carotenoids are also deposited in the flesh (Schiedt et al., 1985). Since the carotenoids are later mobilized from the flesh and transferred to the reproductive organs and eggs, their consummation (algae) may as well act upon the vitamin A1/A2 balance.
The preference of female guppies for a characteristic orange colouration in males determines their mating choice. Moreover, the characteristic orange colouration is regulated by the amount of consumed carotenoids by the males (Deere et al., 2011). In cichlids, a female preference for yellow to red over blue and green dots was observed, which agrees with the appeal of male sexual signals through egg-spots (Egger et al., 2011). The authors suggested that the colour preference is also related to a diet preference of carotenoid rich food, like shrimps, algae and fish eggs. A preference for carotenoid enriched diets is also known in sticklebacks. The intensity of the male red breeding colouration reflects the degree of infection with ectoparasites. Females prefer the more intense red over a less saturated colouration due to external parasitieation in less healthy males (Milinski & Bakker, 1990). A female preference for saturated hues in the male colouration was also found in Pundamilia species (cichlids), which was related to heritable mutations of the LWS opsin genes (Seehausen et al., 2008; Spady et al., 2006).
Although colour preference can be explained ultimately (evolutionary) and may even show a correlation with colour sensitivity (Maan et al., 2006), it remains very difficult to understand the proximate (physiological) origin of a colour preference.
Studies on optional colour preference revealed poor affinity with blue in zebrafish (Avdesh et al., 2010) and in a number of marine fishes (Kawamura et al., 2010). Apparently, these fish have a natural preference for medium and long wave light. In a number of species this preference has shown to be adaptive. Only goldfish seem to like blue (Muntz & Cronly-Dillon, 1966), but the experimental interpretation may be wrong and novel investigations are recommended.
In this context, it should be noticed that the assumed colour preferences of the test animals are based on a diversity of used methods. Since the study of colour preference is usually related to optimal rearing results, one must realize that the most avoided colour does not necessarily coincides with the complementary colour of the most frequently approached one and that there may be a range of colour preferences for ambient light (Luchiari & Pirhonen, 2008) near objects (Egger et al., 2011), tank walls and the floor beneath the test animal (Ullmann et al., 2011; Avdesh et al., 2010). This variation is, for instance, demonstrated in the thinlip mullet (Liza ramada) that is most efficiently raised in light-coloured tanks on a dark-coloured diet (El-Sayed & El-Ghobashy, 2010). < click here to go back to PAN-OPTICS & EVOLUTION |
|
|