| |
Long before quantummechanics enlightened the coherece between radiation and matter, people were fascinated by their relation, probably without realizing. What made plants grow? How could animals see? Why is a sun-bath comfortable?
From early days natural pigments like indigo, alizarin and saffron were used to stain dress materials and garments and their synthesis in the nineteenth century became the kick-off of many chemical industries. The property of materials to selectively absorb or reflect visual light makes our world quite colourful. And this is only because our eyes can discriminate between different wavelengths of the visual spectrum. The latter is due not only to selective absorption but also to the transformation of absorbed light energy into other forms of energy. A property that is not restricted to organic chemicals!
Radiation is mainly determined by its frequency. Low frequencies like radio waves seem harmless to living organisms. In contrast, high frequencies of cosmic radiation and radioactivety are deadly. Even X-rays and UV can do a lot of damage to the living cell.
Radiation damages polymers
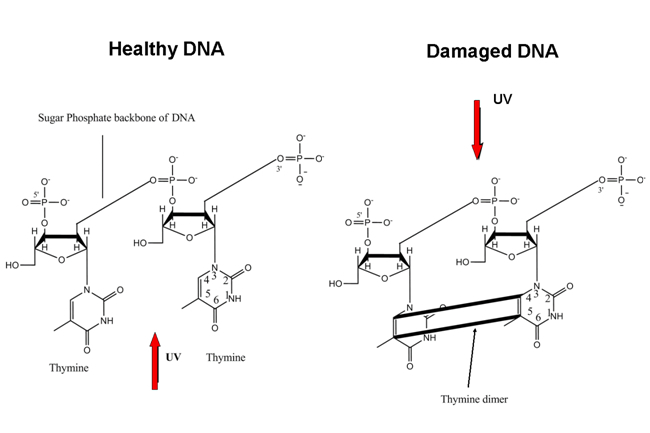
Every living cell contains nucleic acids, like DNA, which are long polymers of nucleotides. There are only a few different nucleotides and their sequence determines the genetic code, i.e. the necessary information for a cell to function properly. The nucleotides are increasingly damaged by the higher frequency radiations and so is the genetic code. As a consequence, the cell dies or starts uninhibited multiplication: it becomes a tumor. The unchecked growth of malignant tumors that use increasing amounts of a body’s nutrients and energy, is known as cancer. Clearly, life on earth must be protected against high frequency radiation coming from the sun. This has sufficiently been realized in the past, when the oxygen in the atmosphere was partly transformed into ozon by the absorption of high frequency radiation, which is still maintained today (although we must be careful with pollution).
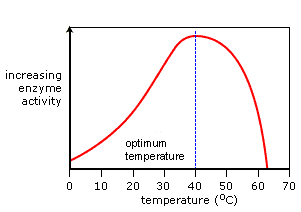
On the other side of the spectrum, microwaves and far IR are perceptible as heat, which is – within limits – essential to living processes. Life in general is possible along a heat-range of liquid water (0 – 100 degrees C) and in most living organisms, proteins start to coagulate above 42oC. Proteins are polymers of amino acids and many proteins are enzymes. Most chemical reactions within the living cell progress smoothly because of the catalystic properties of enzymes. These enzymes are disintegrated by heatwaves: the higher the temperature, the faster the disintegration.
As long as they are not overheated, a sunbath is quite comfortable and to many organisms even necessary, on the condition that the body is protected against too high a level of UV.
Technical applications
Some materials absorb radiation that changes the energy content of the atoms: electrons become excitated, i.e. they increase their distance towards the nucleus. Thus, in some ionic bonds like bromic silver (AgBr), the electron can swich from the bromine ion towards the silver ion and create silver atoms wich are dark-coloured. The principle was discovered and developed in France in the early 19th century and became the staring point of photography.
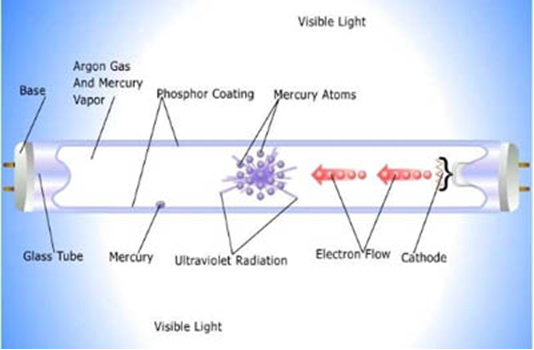
In elements like mercury (Hg) and phosphorus (P), the excitated electrons emit the absorbed energy again, often at a different wavelength. The principle is used in fluorescent lightning, where electrons produce UV that is transformed into visible light.
More recent investigations have focused on the industrial development of radiation-induced adhesive materials. In UV-technology, for instance, invisible ultra-violet radiation is used to quicken the drying of especially composed glue or paint. The technique is also used to accelerate the drying of fillingmaterial and sealing of teeth in dental care. The materials contain a chemical that is activated by the UV radiation and starts a polymerization process. In this way the material is dried and hardened within seconds.
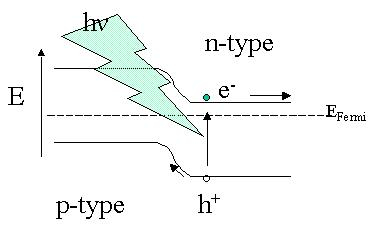
The solar cell converts radiation energy into electricity by means of the photoelectric effect of a semiconducting material like silicon. Quanta of radiation energy are absorbed by the material and excitate electrons, allowing these so called photoelectrons to flow through the material to produce electricity. The most common type of solar cell is basically a large junction between two semiconductors pressed together, each of which has a different kind of electric flow (P- and N-type). N-type silicon has been doped with phosphorus gas to turn it into a material that contains extra electrons that it will release easily. P-type silicon is doped with boron gas to turn it into a material that contains holes that accept a free electron easily. Although "n" and "p" imply negative and positive, n-type and p-type silicon are in an "in-between" stage that has the inclination to readily become more negative or positive.
Photosynthesis
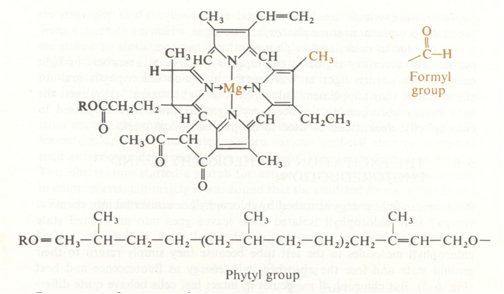
Matter’s capacity to transform radiation energy into an electron flow is probably one of nature’s most fundamental ways to actualize life on earth. Various pigments like (bacterio)chlorophyll, carotenes, xantophyll, phicobiliproteins and others, which all have a specific absorption spectrum, enable plants and some bacteria to transform the absorbed light energy into chemical energy, which in turn is then used to reduce carbon dioxide to form glucose (Lehninger, 1971). In general the hydrogen donor (electron donor) is water or, on some occasions, another reducing agent like hydrogen sulphide or isopropanol, resulting in the generalized equation
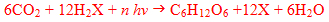
where H2X is the hydrogen donor and X its dehydrogenated form. n hv represents the neceassary amount of light energy. In photosynthesis (or carbon assimilation) the electrons are removed from H2O and transferred ‘uphill’ to CO2, a process that requires energy.
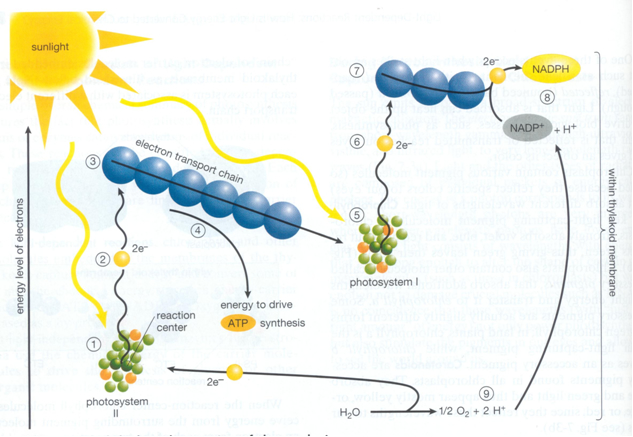
The light-absorbing pigment goes into an excited state when it is illuminated and supported by electron-carrier enzymes in the intact chloroplast, the high-energy electrons leave the excited chlorophyll molecule. During this process the electrons are passed on to NADP molecules (nicotinamide adenine dinucleotide phosphate) which are then reduced to NADPH. Next to this noncyclic process, the electron flow is used to form ATP (adenosine tri-phosphate) by photophosphorylation of ADP while the electrons return to the chlorophyll (cyclic photophosphorylation).
In the eukaryotic cells containing chloroplasts the two major light-absorbing pigments are chlorophyll a and chlorophyll b that generate photosystem I and photosystem II respectively.
Removal of electrons from water causes ultimately the formation of molecular oxygen
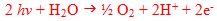
and the overall equation may be written as
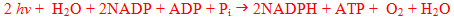
where the molecules of H2O that enter are the electron donors for reduction of NADP and the source of molecular oxygen; the H2O molecule appearing on the right-hand side is that extracted from phosphate and ADP to yield ATP. Green plants containing chlorophyll a and b are oxygen-evolving photosynthetic organisms in contrast to organisms containing bacteriochlorophyll. This is subject to opposing ideas on the evolution of photosynthesis (Lockhart et al., 1996).
The reduced NADP and the energetic molecules ATP are then used to produce glucose out of a number of carbonoxide molecules. This occurs in a complex sequence of enzyme-catalyzed reactions called the Calvin-Benson cycle and is light-independent.
Photosynthesis does not always require chlorophyll. Some algae and bacteria may have a different system based on retinal, which also could convert light to chemical energy (Ebrey, 2002)
Photoreception
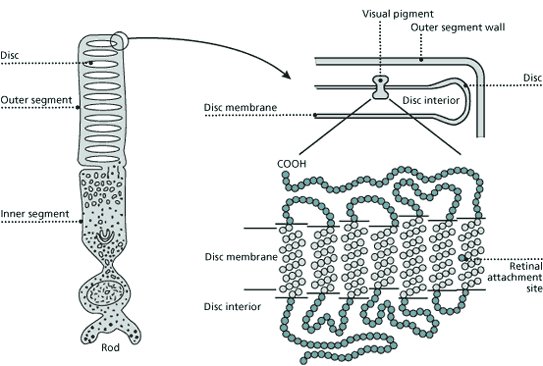
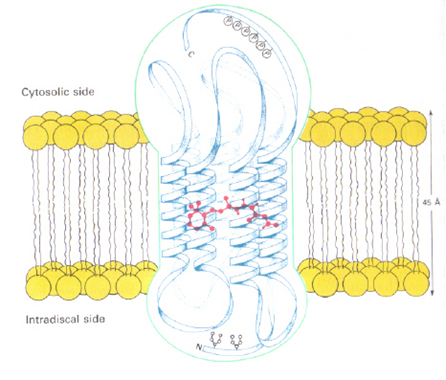
In virtually all multicellar animals vision is generated by photoreceptors, using a light-sensitive pigment called retinal (the chromophore) that is derived from vitamin A, and that is bound to a protein called opsin. The photoreceptors (rods and cones) usually contain rhodopsin (11-cis retinal bound to an opsin). Organisms like many fresh-water fish living in a turbid environment where the light is shifted to the longer wavelengths, have or porphyropsin (11-cis 3-dehydroretinal bound to an opsin) instead of rhodopsin, or a mixture of both. Porphyropsins have absorption spectra displaced to longer wavelengths (Bridges, 1990).
There may be only one chromophore (retinal), but because the opsins are a little different, the way the retinal is attached to an opsin has a great deal of affect on the wavelength of its maximum absorption. There are various theories to explain why the same molecule absorbs light at different wavelengths. One theory is related to how the protein arranges itself around the retinal and to the bend in the carbon structure of retinal. When the structure is bent, the carbons are not in the same plane. The more bent the structure the more blue-shifted the light absorption is going to be. Another hypothesis relates to the point at which retinal attaches to the opsin. At the attachment-point there is a nitrogen molecule with a positive charge. Because the positive charge requires a negative charge, the location of that negative charge is very crucial. The retinal becomes more planar in the active site as the distance between the positive and negative charges increases. Therefore, it is hypothesized that the negative charge and positive charge are very close in blue rhodopsin whereas they are far apart in the red rhodopsin.
There is evidence that the spectral sensitivity of the opsins depends on the amino acid sequence within the protein. Furthermore, the difference in phototransduction proteins between rods and cones is probably responsible for their sensitivity to light (Hisatomi et al., 2002)
The general process of photoreception can be described as follows. When a light beam hits 11-cis retinal, it changes into all-trans retinal which causes a conformational change in the photopigment, and forces the opsin to bind to a G-protein. This protein then releases a subunit of its molecular structure, the alpha segment. This subunit attaches itself to another protein leading to a cascade of events resulting in closure of Ca2+-channels within the receptor-cell. The resulting gradient in ion concentrations between both sides of the cell-membrane initiate an electrical impulse that leads to the release of a neurotransmitter near the synapse. The neurotransmitter is picked up by the next neuron and the signal is finally transmitted to the brain where it may contribute to a visual image.
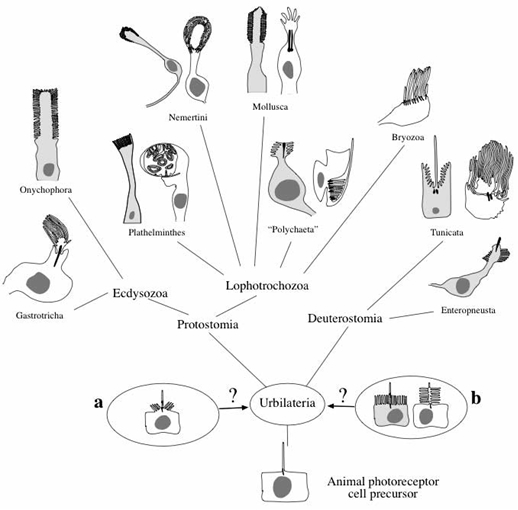
The formation of the eyes of multicellular animals is regulated by a common homeobox gene, a pax6 homolog. Whether rhabdomeric (arthropods) or ciliary (vertebrates), all photoreceptors use a light-sensitive pigment derived from vitamin A, and this pigment is bound to a protein. Light activates the opsin by causing a conformation change in the photopigment, and the opsin then binds to a G-protein, a common and versatile molecule used in many signal transduction cascades. These similarities suggest that all eyes have a common evolutionary ancestor. (http://scienceblogs.com/pharyngula/2006/09/rhabdomeric_and_ciliary_eyes.php).
< click here to go back to PAN-OPTICS & EVOLUTION |
|
|